1. Exercise that involves stretching a muscle while active cause microscopic areas of damage, delayed onset muscle soreness, and adaptation to withstand subsequent similar exercise.
2. Longer muscle lengths are associated with greater damage, and recent animal experiments show that it is the length relative to optimum that determines the damage.
3. In humans walking down stairs, taking two at a time increases the length of the muscle during the lengthening and increases the delayed onset muscle soreness.
4. The observed pattern of damage is consistent with explanations based on sarcomere length instabilities
5. The pattern of adaptation is consistent with the number of sarcomeres in series in a muscle being modulated by exercise, especially the range of muscle lengths over which eccentric exercise regularly occurs.
When muscle is stretched during active tension generation, the event is commonly, though not intuitively, described as an eccentric contraction. Other terms include pliometric contraction, or active stretch. The muscle is being forcibly lengthened while trying to shorten. Energetically the muscle is absorbing work, not performing it. In common terms, the muscle is being used as a brake, not a motor.
Eccentric contraction is an important function of muscle, occurring during activities ranging from lowering a load to walking down hill. It is present in many forms of exercise, such as running, particularly when down hill is included, horse-riding and skiing, where the action could be better described as shock absorbing rather than braking. Eccentric contractions seldom occur in cycling, rowing or swimming.
It is a long standing observation that "unaccustomed" eccentric exercise can lead to stiff and tender muscles next day, known as Delayed Onset Muscle Soreness, or DOMS1. It has become apparent in recent years that "unaccustomed" can mean at unaccustomed length, as well as involving an unaccustomed number of repetitions or unaccustomed forces2,3. This is a key observation that will be reinforced later. DOMS is accompanied by microscopic muscle damage, with multiple areas of damage scattered throughout the muscle, but each confined to a single fibre4. The maximum force is also reduced, the dependence of force on stimulation rate is changed5, and the optimum length is immediately shifted to longer lengths6. In some experiments, usually in frog fibres, the tension at long length has been shown to increase, making the changes in activation unable to fully account for the shift in optimum length6.
Perhaps most importantly, eccentric exercise produces a rapid adaptation, so that a second similar bout of exercise produces substantially less soreness and injury7.
The popping sarcomere hypothesis8 states that stretch induced muscle damage results from very non-uniform lengthening of sarcomeres when active muscle is stretched beyond optimum length. If sarcomeres are beyond optimum length, then the longest sarcomeres will be the weakest, and so will be stretched more rapidly than the others, and so become weaker, until rising passive tension compensates for falling active tension. For at least some muscles, this corresponds to lengths beyond filament overlap. (The situation in other muscles is unclear, depending on the origins of the resting tension.) As the weakest sarcomeres are not at the same point along each myofibril, this non-uniform lengthening leads to shearing of myofibrils, exposing membranes, especially t-tubules, to large deformations. This is thought to lead to loss of calcium ion homeostasis, and hence damage, either through tearing of membranes or opening of stretch activated channels9. It is postulated8 that the adaptation to eccentric exercise consists of increasing the number of sarcomeres in series, so that a given muscle length corresponds to a shorter sarcomere length. In particular, the adapted muscle confines eccentric activity to muscle lengths less than optimum.
For a simple muscle where all fibres have the same number of sarcomeres in series, the optimum length for tension generation during maximal activation will be given by the length of tendons, plus the product of the number of sarcomeres in series and the optimum length of a sarcomere. For a muscle containing fibres with a range of the number of sarcomeres in series, the above relation will approximately hold if the number of sarcomeres in series is replaced with the average number of sarcomeres in series. As the optimum length of a sarcomere is fixed, and tendons are slow to remodel, shifts in the optimum length with exercise that occur within a few days can be confidently assigned to changes in the number of sarcomeres in series.
In the popping sarcomere hypothesis, the optimum muscle length takes on three very important roles. In the first, optimum length becomes a prime determinant of the susceptibility; damage is predicted to occur only when sarcomeres are used beyond optimum length. In the second, the immediate shift in optimum length after eccentric exercise is a measure of damage, which is largely specific to eccentric exercise damage, and independent of fatigue or damage that involves fibres ceasing to contract, both of which contribute to force drop. Here it is proposed that the immediate shift indicates the number of overstretched sarcomeres, an early stage in the process leading to DOMS. Experimentally, the shift in optimum is closely correlated with the fall in tension, but typically shows less scatter2. In its third role, the position of the optimum is a measure of adaptation. A muscle is expected to be protected from injury if the optimum length is near to or beyond the maximum length at which the muscle undergoes eccentric contraction.
The importance of long length in determining damage has been clearly demonstrated in frog sartorius muscles, rat vastus intermedius, and cat gastrocnemius. Furthermore it has been shown that damage to an individual motor unit within a muscle undergoing eccentric exercise depends on that motor unit's optimum length10.
A recent unpublished experiment from our laboratories also showed a dependence of damage on length for human knee extensor muscles. When we walk down stairs, the knee extensors of the supporting leg undergo an eccentric contraction as they support the body as it is lowered to place the extended leg on to the next step. This does not normally cause muscle soreness, as it is an everyday exercise. Descending the same distance two steps at a time causes the muscles to undergo eccentric contractions at an unaccustomed longer length, while still generating the same forces and absorbing the same amount of energy.
Twenty one students walked down ten flights of 24 stairs with instructions to land on their heels rather than their toes. For alternate flights, they stepped down one step with leg A leading, and then brought leg B alongside, so that leg B absorbed all the energy of descending one flight in 24 eccentric contractions at short length, while the knee extensors of leg A were essentially inactive. For the other flights, they stepped down two steps with leg B leading, and then brought leg A alongside, so that A was the supporting leg that underwent eccentric contraction at long length. The same amount of energy was absorbed as for the other flights, but spread over only 12 eccentric contractions extending to longer muscle length. The relations between left and right, dominant and non-dominant, and A and B were randomized.
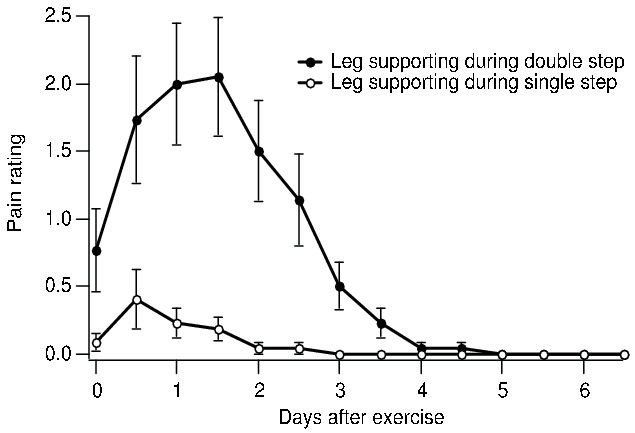
Subjects rated soreness in their left and right quadriceps muscles twice daily for the next week on a scale of 0 to 10, representing no pain and extreme pain respectively. Most experienced only mild pain. Figure 1 shows that the mean pain rating for leg A, the support leg during the double step, reached 2 between 24 and 48 hours, but leg B only reached 0.4. A General Linear Model of the 616 pain ratings was carried out for the following factors. The time of measurement was tested as a discrete variable, due to the rise and fall of pain with time, and reached p<0.0001. The time course is shown in Figure 1. The comparison between the A and B legs, that is whether the leg had been supporting the body during single or double stepping had p<0.0001, with the leg that was stretched to longer lengths showing more pain. The length of the subjects' legs, measured as the height of the iliac crest above the floor was significant as a linear factor with p<0.003, with taller people experiencing less pain, presumably because the fixed step requires smaller angles of flexion for taller people. Subjects also graded their regular participation in eccentric and concentric exercise on a five point scale. From these, an exercise factor was calculated as the difference between the index of participation in eccentrically biased exercise and the index for concentrically biased exercise. This showed p=0.0001, with high scores showing less damage. Gender had p<0.0001 when leg length was removed as a factor, but this became p<0.02 when leg length was included, suggestion that gender primarily acted through height. Weight was not a significant continuous factor. Measurements of optimum length were not made.
Figure 1. Mean with SEM of pain ratings of knee extensor muscles after walking down stairs supported by one leg for five flights taken one step an a time, and the other leg for five flights taken two steps at a time. The larger steps stretched the muscles to longer lengths and produced more soreness.
These results are all in accord with the hypothesis that damage only occurs when muscle is actively stretched beyond its optimum length, and that muscles adjust their optimum length by adjusting the number of sarcomeres so that active stretch normally occurs only at lengths less than optimum.
The idea that adaptation to eccentric exercise consists of adding sarcomeres in series in fibres has a number of supporting observations. It ties in closely with the observations that damage depends critically on the range of lengths of the stretch compared to optimum. It also explains why biopsy studies have been unable to show differences with adaptation, as the sarcomeres are unchanged. It is consistent with previous observations that the number of sarcomeres in series is capable of relatively rapid change11, though the mechanisms are yet to be fully elucidated. It also gives a reason for the reversal of training. Extra sarcomeres in series increase the energy consumption for isometric force development. This provides an incentive to shed un-needed sarcomeres in the interests of efficiency of tension generation. Extensive exercise involving only concentric contractions would be expected to increase this, supporting the anecdotal observation that "couch potatoes" are less prone to eccentric exercise damage to knee extensors than regular cyclists. In this view, the number of sarcomeres is continuously modulated up or down to maximise efficiency while avoiding damage during "normal" activity. Investigating the mechanisms of the adaptation is beyond our expertise.
Direct evidence for sarcomere number modulation has come from rat vastus intermedius muscles, the postural knee extensors12. Rats were trained by running on an climbing or descending treadmill for about 20 mins per day for five days, in a protocol that had been previously shown to cause damage to these muscle in the descending group but not the climbing group13. The muscles were fixed and then digested in acid. Intact fibres were identified in serial dilutions of the digested muscle, and the number of sarcomeres estimated from fibre length and sarcomere length, measured by laser diffraction. From these the number of sarcomeres in fibres was calculated. All groups had quite broad distributions of sarcomere numbers, but the means were significantly different between training groups. The descending trained animals had the largest sarcomere count, the climbing trained rats had the smallest counts, and sedentary rats had intermediate counts, though closer to the climbing group. This could be interpreted as the response to concentric exercise being either smaller or slower than the response to eccentric exercise.
In another series of experiments14, vastus intermedius muscles of treadmill trained rats were tested mechanically while still in situ, that is attached to the bones, but with all other muscles about the knee joint removed. This avoids introducing uncertainties into muscle length. In descending trained rats the knee angle for optimum torque generation corresponded to longer muscle lengths than in climbing trained rats, and the muscles of descending trained rats suffered less damage from an acute bout of eccentric contractions over the same range of knee angles.
In humans the hamstring muscles are of particular interest, because they are used relatively rarely for eccentric contractions, and because they are subject to muscle tears, which are thought to occur during eccentric contractions in activities such as sprinting where they act as brakes on the forward swing of the leg, particularly the lower leg. They are also a convenient muscle to use experimentally, as passive tension about the knee is small over most of the anatomical range. The optimum knee angle for torque generation can be reliably measured by isokinetic dynamometry, where an angle torque curve is measured during maximum voluntary contraction with constant velocity shortening. Doing a number of cycles and averaging improves the reliability.
Using this measure before and after a series of eccentric exercises produced a significant shift of about 7� in optimum knee angle for torque generation15. The exercise was a series of "hamstring lowers", where the subject knelt on a pad with ankles restrained and hips straight, and leaned slowly forward from the knees as far as possible before collapsing. This led to muscle soreness in the hamstring group. Some subjects repeated the exercise ten days later, and suffered much less soreness. Again eccentric exercise, DOMS, increased optimum length and adaptation all occurred together.
A less well investigated consequence of the hypothesis is that the unloaded shortening velocity should increase with eccentric training, as the unloaded shortening velocity of a fibre is the sum of the velocities of the sarcomeres. Some indications of this have been reported16, though the long time course makes it difficult to rule out fibre type change as an alternative explanation.
It has been noted in tetanically stimulated animal muscles with multiple fibre types that the damaged fibres are largely fast twitch fibres17. In sub-maximal voluntary activity the opposite is true13, presumably because the fast twitch fibres are not likely to be activated. Hence the fast fibres can be seen as being susceptible in tetanic contractions because they are not recruited, and hence not trained, during normal submaximal activity. If training consists of increasing the number of sarcomeres, then slow motor units should have longer optima than fast units, or more precisely, the most susceptible units should have the shortest optima and be fast type. This was tested for cat gastrocnemius motor units10. It was found that all motor units with optimum length less than the whole muscle optimum had a time to peak twitch of less that 50msec, that is all slow motor units had long optima. Interestingly, a small number of fast twitch units had long optimum lengths, suggesting that they have adapted to, and hence were subject to, regular eccentric exercise. However as the experimental observation was that all damaged fibres were fast, not all fast fibres were damaged, the existence of units that are both fast and well adapted is not inconsistent.
Different optimum lengths may be due to different tendon lengths as well as different numbers of sarcomeres. Where a relatively rapid (hours to days) shift of optimum is shown, it is more likely that sarcomeres are involved than tendon. This is less clear with experiments such as the above comparison of motor units, where the adaptation has taken place over a lifetime, so that tendon adaptations cannot be ruled out. However, the basic hypothesis that muscle adapts optimum length to avoid eccentric contractions beyond optimum is not affected by this complication.
It is possible that adaptation of optimum length does occur, but that other adaptation mechanisms are also important. This was tested by training more rats as described above, either climbing or descending. All animals were then anaesthetised, and the vastus intermedius subjected to a series of 20 eccentric contractions. All stretches had an amplitude of 27� of knee angle, but beginning at different knee angles. In some animals stretch began at the measured optimum angle, while in others it began from 90� of knee angle3. One way of analysing these data is shown in Figure 2. Damage has been quantified by the immediate shift in optimum angle, but the tension decrease shows similar results. When the damage is plotted against the absolute knee angle from which the eccentric contractions began, the climbing and descending trained muscles clearly fall on two distinct lines, confirming both the existence of a training effect and the dependence of damage on muscle length. When the same damage data are plotted against the angle relative to optimum from which the acute eccentric contractions began, the two groups fall on the same line. Damage still depends on sarcomere length, but not independently on training. Statistical analysis of the data confirmed these conclusions. If training group (discrete) and absolute knee angle (continuous) were used as factors in a General Linear Model, group was highly significant. If absolute knee angle was replaced by relative knee angle, group was no longer significant. This indicated that effect of training attributable to mechanisms other than the shift in optimum length was not statistically significant.
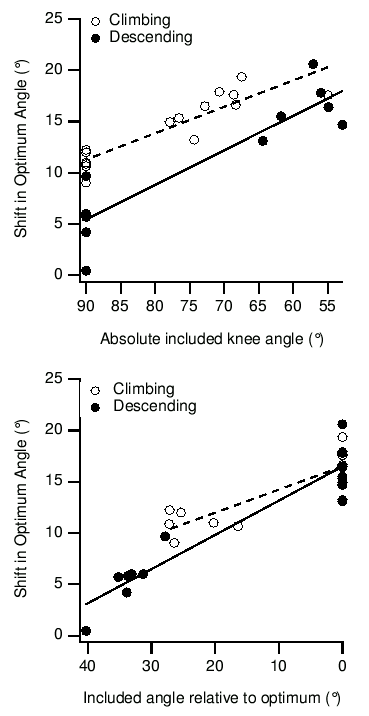
Figure 2. The shift in optimum angle after acute eccentric contractions from various initial lengths, in climbing and descending trained rats. In the upper panel, data are plotted against the absolute knee angle from which stretches began. The muscles stretched from 90� are on the left, and those stretched from optimum length were generally longer and more damaged. Most importantly, the climbing and descending animals are clearly separated. In the lower panel the same data are plotted against angle relative to optimum. In this case the muscles stretched from optimum all appear at zero, and those stretched from 90� are generally stretched from less than optimum and damaged less. In this case, the climbing and descending trained animals are not separated.
These ideas and observations have been useful in directing current research into eccentric exercise, such as examination of the role of transverse tubules9 and in muscle injury prevention18.
There is now an extensive body of evidence that damage from eccentric exercise is strongly dependent on the sarcomere lengths over which the stretching occurs. Adaptation in a number of preparations has been shown to be accompanied by a shift in optimum length, and the shift has been shown to account for the major portion of the adaptation. All of this provides evidence that damage occurs when sarcomeres are beyond optimum, the central prediction of the popping sarcomere hypothesis. This is true whether the stretch progresses to damage by tearing of structures or by opening of stretch activated channels.
1. Hough T. Ergographic studies in muscular soreness. Am. J. Physiol. 1902;7:76-92.
2. Talbot JA, Morgan DL. The effects of stretch parameters on eccentric exercise induced damage to toad skeletal muscle. J. Musc. Res. Cell. Mot. 1998;19:237-245.
3. Morgan DL, Talbot JA. The Addition of Sarcomeres in Series is the Main Protective Mechanism Following Eccentric Exercise. J. Mech. Med. Biol. 2002;2(3 & 4):421-431.
4. Ebbeling CB, Clarkson PM. Exercise-induced muscle damage and adaptation. Sports Med 1989;7:207-234.
5. Edwards RHT, Hill DK, Jones DA, Merton PA. Fatigue of long duration in human skeletal muscle after exercise. J. Physiol. (Lond.) 1977;272:769-778.
6. Morgan DL, Claflin DR, Julian FJ. The effects of repeated active stretches on tension generation and myoplasmic calcium in frog single fibres. J. Physiol. (Lond.) 1996;497(3):665-674.
7. Newham DJ, Jones DA, Clarkson PM. Repeated high-force eccentric exercise: effects on muscle pain and damage. J. Appl. Physiol. 1987;63(4):1381-1386.
8. Morgan DL. New insights into the behavior of muscle during active lengthening. Biophys. J. 1990;57(Feb):209-221.
9. Yeung EW, Balnave CD, Ballard HJ, Bourreau J-P, Allen DG. Development of T-tubular vacuoles in eccentrically damaged mouse muscle fibres. J. Physiol. (Lond.) 2002;540(2):581-592.
10. Brockett C, Morgan DL, Gregory JE, Proske U. Damage to different motor units from active lengthening of the medial gastrocnemius muscle of the cat. J. Appl. Physiol. 2002;92:1104-1110.
11. Goldspink G. Malleability of the motor system: a comparative approach. J. Exp. Biol. 1985;115:375-391.
12. Lynn R, Morgan DL. Decline running produces more sarcomeres in rat vastus intermedius muscle fibres than incline running. J. Appl. Physiol. 1994;77(5):1439-1444.
13. Armstrong RB, Ogilvie RW, Schwane JA. Eccentric exercise induced injury to rat skeletal muscle. J. Appl. Physiol. 1983;54(1):80-93.
14. Lynn R, Talbot JA, Morgan DL. Differences in rat skeletal muscles after incline and decline running. J. Appl. Physiol. 1998;85(1):98-104.
15. Brockett C, Morgan DL, Proske U. Human hamstring muscles adapt to damage from eccentric exercise by changing optimum lengths. Med. Sci. Sports Exercise 2001;33(5):783-790.
16. Fridén J, Seger J, Sjöström M, Ekblom B. Adaptive response in human skeletal muscle subjected to prolonged eccentric training. Int. J. Sports Med. 1983;4:177-183.
17. Lieber RL, Fridén J. Selective damage of fast glycolytic fibres with eccentric contraction of the rabbit tibialis anterior. Acta Physiol. Scand. 1988;133:587-588.
18. Brockett C, Morgan DL, Proske U. Predicting hamstring strain injury in elite athletes. Med. Sci. Sports Exercise 2004;36(3):379-387.