1. The renal nerves constrict the renal vasculature causing decreases in renal blood flow (RBF) and glomerular filtration rate (GFR). Whether renal haemodynamics are influenced by changes in renal nerve activity within the physiological range is a matter of debate.
2. We have identified two morphologically distinct populations of nerves within the kidney, which are differentially distributed to the renal afferent and efferent arterioles. TYPE I nerves almost exclusively innervate the afferent arteriole whereas TYPE II nerves are distributed equally on the afferent and efferent arterioles. We have also demonstrated that TYPE II nerves are immuno-reactive for neuropeptide Y while TYPE I nerves are not.
3. This led us to hypothesise that in the kidney, distinct populations of nerves innervate specific effector tissues and that these nerves may be selectively activated, setting the basis for the differential neural control of GFR. In physiological studies, we demonstrated that differential changes in glomerular capillary pressure occurred in response to graded reflex activation of the renal nerves, compatible with our hypothesis.
4. Thus, sympathetic outflow may be capable of selectively increasing or decreasing glomerular capillary pressure and hence GFR by differentially activating separate populations of renal nerves. This has important implications for our understanding of the neural control of body fluid balance in health and disease.
Opinion as to the importance of the renal nerves in controlling RBF and GFR has risen and fallen over the last 150 years. In the first study to demonstrate a role for the renal nerves in the control of renal function, Claude Bernard in 18591, transected the renal nerves and noted a marked diuresis, which he attributed to an increase in RBF. This and similar studies in the years following, demonstrating the phenomenon of denervation diuresis, dominated the understanding of the neural control of renal function (see 2). During this period the renal nerves were thought to exert a profound effect on the regulation of RBF and GFR.
Yet 80 years later, opinion had swung full circle, when Homer Smith dismissed the renal nerves in his landmark book The Physiology of the Kidney 2 as having little importance in the control of renal function except in cases of severe stress. Smith damningly wrote of Bernard’s study, stating that “His conclusion is admittedly correct, but his experiment was unfortunate in two respects.”2. In the first instance, the development of clearance techniques to measure RBF and GFR demonstrated that changes in urine flow rate do not reflect changes in RBF. Secondly the anaesthesia and surgical stress to which the animals were subjected resulted in elevated basal levels of renal nerve activity. Bernard’s finding of increased urine flow, probably was associated with an increase in RBF, but was due to the release of the kidney from the stress-induced increase in renal nerve activity. Smith’s own studies, made painlessly in conscious and unstressed animals, failed to demonstrate any change in RBF or GFR following renal denervation. It was concluded that kidney function was not dependent on tonic renal sympathetic activity2. Soon after the first kidney transplantations were performed, the apparent lack of long-term effects on body fluid balance was taken as confirmation of the independence from nervous system control of renal vascular and tubular function3.
However, in the 1970’s there was a resurgence of interest in the neural control of renal function, sparked by quantitative analysis of the distribution and density of neuroeffector junctions in the kidney4 and appreciation that transplanted kidneys rapidly re-innervate5.
Today it is widely accepted that changes in renal sympathetic nerve activity (RSNA) play a significant role in controlling body fluid homeostasis during normal daily activity and in the pathophysiology of many clinical conditions6-8. Whether this is primarily due to changes in renin release and tubular reabsorption, or also involves changes in RBF and GFR, is debated (see 9,10).
In this review evidence is considered which supports the hypothesis that different populations of renal nerves selectively affect the afferent and efferent arterioles thereby allowing differential control of glomerular capillary pressure and hence single nephron glomerular filtration rate (SNGFR).
A brief outline of the physiological basis of the control of glomerular ultrafiltration is necessary to understand how the renal nerves might contribute to its control. More detailed accounts can be found elsewhere (see 11).
The primary force driving SNGFR is glomerular capillary pressure. Precise control of this pressure is important as significant falls in glomerular capillary pressure can lead to acute renal failure, whereas increased glomerular capillary pressure causes irreversible glomerular damage that leads to nephron loss and chronic renal disease12.
The unique arrangement in the kidney of two resistance vessels in series, the afferent and efferent arterioles, allows fine regulation of pressure in the glomerular capillaries13,14. RBF only becomes an important factor in determining SNGFR under conditions of filtration equilibrium, which is not the normal physiological state13,14. Thus, glomerular capillary pressure and therefore SNGFR will increase if the pre (afferent) to post (efferent) glomerular resistance ratio decreases and decrease if this resistance ratio increases.
Importantly, for the majority of glomeruli the resting diameter of the efferent arteriole is smaller than the afferent arteriole15-18. Since resistance is inversely proportional to the fourth power of the radius this explains, in part, how this relatively small, sparsely muscled vessel can counterbalance the effects of constriction of the bigger, more muscular afferent arteriole (see 17 for a more detailed explanation). For juxtamedullary glomeruli - those 10% of glomeruli, whose efferent arterioles descend into the medulla to form the vasa recta - the situation is different since these efferent arterioles are as large if not larger than their afferent counterparts. Therefore the control of glomerular capillary pressure may well be different in juxtamedullary nephrons.
SNGFR can also be influenced by alterations in the glomerular capillary ultrafiltration coefficient (Kf), which represents the product of the glomerular capillary surface area available for filtration and hydraulic conductivity. Kf has been shown to decrease in response to a number of vasoactive stimuli, though the mechanisms are not well understood (see 11).
The kidney receives an extensive sympathetic innervation. While it is generally agreed that all the major structural elements of the kidney are innervated, including vascular smooth muscle cells, renin secreting cells, mesangium and tubules (proximal, distal and loop of Henle)19,20, the relative density of the innervation of each tissue type has been disputed20. Whether functionally specific or non-specific renal sympathetic nerve fibres innervate the effector cells has also been questioned. Barajas concluded that the sympathetic innervation of the kidney was diffuse and non-specific, based on observations that each sympathetic axon made contact with multiple effector tissues (see 19). These studies powerfully influenced how the nerves were thought to control renal function (see 21). However, the definition of a neuroeffector contact in Barajas’s studies is now considered very broad. Varicosities that were separated from the effector cell by up to 300 nm and in which two layers of basal lamina were present were included (see 19).
Our definition of a neuroeffector junction is much more specific20,22. The varicosities along an axon can be divided into contacting and non-contacting. Contacting varicosities form specialized junctions with the effector cell; neuroeffector junctions. The vesicles are organized within the varicosity in clusters associated with the region on the membrane that is in contact with the effector cells. At the point of contact, the varicosity and effector cell are separated by a gap that is less than 100 nm20,22. Consequently, contacting varicosities release neurotransmitters directly onto junctional receptors rather than relying on diffusion of the transmitter to receptors across the surface of the smooth muscle cell membrane20,22. Based on this definition of a neuroeffector junction, we re-examined the innervation of the kidney and drew vastly different conclusions to those drawn from the studies of Barajas (see 19).
Using three dimensional reconstruction ultrastructural analysis of serial thin sections to examine the innervation of the juxtaglomerular region, we identified two ultrastructurally distinct types of sympathetic axons23. TYPE I axons were large in diameter with atypical varicosities and TYPE II axons resembled those innervating blood vessels in other organs, with typical fusiform varicosities23. These axon types were identified in rats and rabbits23. At the time the functional significance of two axon types was unknown, though conduction velocities would be expected to be different. Later, in support of our study, another study demonstrated that there was a bimodal distribution in the diameters of the renal nerves with different conduction velocities 24.
Next we described the distribution and density of neuroeffector junctions made by these two types of axons20. Several important findings were made: (i) The sympathetic axons were located in regions adjacent to the renal vasculature and therefore primarily the arterial vessels were innervated. However, the majority of tubular tissue in the cortex was not innervated. (ii) The afferent arteriole was the most densely innervated tissue. The afferent arterioles were 3 times more densely innervated than the efferent arterioles (Fig. 1 & 2). (iii) There was little evidence for individual axons innervating more than one effector cell type. (iv) Most significantly, it was shown that TYPE I axon varicosities made contact almost exclusively with afferent arterioles whereas TYPE II axons innervated both arterioles at similar densities (Fig. 1 & 2).
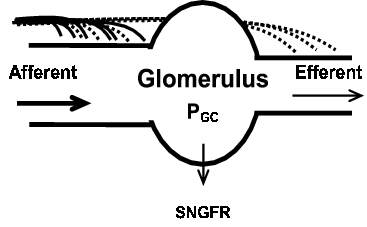
Figure 1. Diagram representing the relative TYPE I (solid line) and TYPE II (dashed line) axon innervation density on afferent and efferent arterioles. The afferent arteriole is 3 times more densely innervated than the efferent arteriole. TYPE I axons (solid lines) almost exclusively innervate the afferent arteriole. TYPE II axons (dotted lines) are equally distributed on the afferent and efferent arterioles. (PGC, glomerular capillary pressure. SNGFR, single nephron glomerular filtration rate).
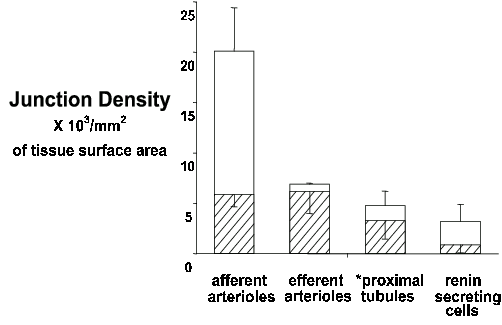
Figure 2. Stacked bar graph of the density of neuroeffector junctions of TYPE I (open bar) and TYPE II (hatched bar) axons on the afferent arterioles, efferent arterioles, proximal tubules (*only those adjacent to the afferent arterioles were innervated) and renin secreting cells. The combined bar equals the total junction density on each effector tissue.
This finding was of great potential significance, and was also recognized as such by others, being rapidly incorporated into standard textbooks on the kidney25,26. It raised the possibility that TYPE I and II axons originated from different populations of neurons.
The presence of distinct combinations of immuno-histochemically detectable substances can be used to identify populations of nerves serving different functions27,28. On this basis, we have recently shown that neuropeptide Y is located in TYPE II axons whereas TYPE I axons lack neuropeptide Y29. Our findings are in good accord with the study of Reinecke et al.30 who reported that the density of neuropeptide Y positive terminals was very similar on the afferent and efferent arterioles; that is, similar to the distribution of TYPE II axons. This provides further evidence that TYPE I and II axons originate from separate populations of neurons. The search for a neuropepetide specific to TYPE I axons is on going.
On the basis of our morphological and immuno-histochemical evidence, we hypothesised that different patterns of sympathetic outflow to the kidney may evoke selective changes in pre- and post-glomerular vascular resistance to regulate GFR.
We hypothesise, based on the distribution of the TYPE I and TYPE II nerves on the afferent and efferent arterioles that (see Fig. 1 & 3), (i) Selective TYPE I axon activation would result in pre-glomerular vasoconstriction, reduced RBF, a reduction in glomerular capillary pressure and a fall in GFR. (ii) Selective TYPE II axon activation would result in pre- and post-glomerular vasoconstriction and decreased RBF. However, the effect on resistance would be greater on the efferent arteriole, since it is a smaller vessel (Poiseuille’s law), leading to little effect on glomerular capillary pressure resulting in the maintenance of GFR17. (iii) Activation of both TYPE I and II axons would cause a predominant decrease in pre-glomerular vascular resistance due to the greater innervation density of the afferent arteriole. We have pursued this possibility in physiological studies outlined below.
The sympathetic nervous system is capable of producing selective changes in efferent outflow to different organs (see 31-33). Increasing knowledge of central autonomic nervous system organisation, indicates that the output to different sympathetic pre-ganglionic neurons depends on the relative contributions of a wide range of brain nuclei and on the particular pattern of inputs to those nuclei (baroreceptor, chemoreceptor, somatic receptors and inputs from all areas of the brain)31. We are proposing within the kidney, as has been demonstrated in other organs (eg there are at least 3 distinct types of sympathetic neurons in the gut28), that there is further differentiation of the signal such that specific effector tissues may be selectively activated, depending on the nature and severity of the stimulus. In the literature there is limited and conflicting evidence as to whether subpopulations of renal post-ganglionic nerves can selectively regulate different renal functions34,35.
Current views on the neural regulation of renal function rely mainly on data from electrical stimulation studies, or even the effects of simple acute denervation. It has been widely accepted that individual renal nerves innervate multiple tissues (vascular smooth muscle, renin secreting cells and proximal tubules)19 and that renal function is affected entirely by the frequency of their firing, with low to moderate frequencies stimulating increased renin release and sodium reabsorption and only high frequencies stimulating a decrease in renal blood flow and GFR (see 21). According to this view renal sympathetic nerve activity (RSNA) is generally too low to influence renal vascular resistance and glomerular ultrafiltration under normal physiologic conditions21. This does not accord however with a large body of physiological and clinical evidence that suggests that renal hemodynamics are under the control of RSNA during daily events (see 9).
Not surprisingly, since the afferent arteriole is much more densely innervated than the efferent arteriole20, electrical stimulation of the renal nerves results in a predominant increase in pre-glomerular resistance36. This causes glomerular capillary pressure and GFR to decrease, as predicted when both TYPE I and II nerves are fired simultaneously (see Fig 3). These studies therefore shed no light on the possible effects of selective physiological recruitment of different populations of renal nerves on the renal resistance vessels.
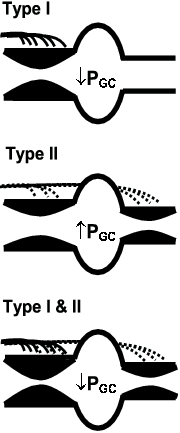
Figure 3. Diagram demonstrating the hypothetical effects of selective activation of TYPE I (upper panel), TYPE II (middle panel) or both TYPE I and II (lower panel) axons innervating the afferent and efferent arteriole, on glomerular capillary pressure (Pgc). See text for explanation.
RSNA varies in both the frequency (reflecting the rhythms of the central generating circuits and baroreflex input) and amplitude of its discharge (reflecting the relative number of activated nerves)37,38. Thus, whereas electrical stimulation activates all nerves simultaneously, it is now evident that relatively few individual nerves are active at rest and that the number of nerves activated during physiological bursts of nerve activity varies widely39. Physiological activation of the renal sympathetic nerves is therefore fundamentally different to electrical stimulation.
A number of studies have examined the kidney’s response to reflex activation of the renal sympathetic nerves. Again not surprisingly, in response to severe increases in RSNA, when many nerves are firing, RBF and GFR decrease (see 21), indicative of TYPE I and/or II nerve firing (see Fig. 3). However, in several studies no change in GFR was reported in response to moderate increases in RSNA40-43. It is quite possible that in these studies subtle changes in pre- and post-glomerular vascular resistances were occurring to maintain glomerular capillary pressure and GFR. However, no study measured glomerular capillary pressure to verify such a conclusion.
At this time, it is not possible to identify individual Type I versus Type II nerves in vivo, and thus selectively record or stimulate these neurons. However, we do have tools whereby we can determine whether the pattern of changes in pre-and post-glomerular vascular resistance in response to reflex stimulation of RSNA is presumptive of TYPE I nerve recruitment, TYPE II nerve recruitment or both. In vivo micropuncture is a challenging and time consuming procedure, but it is the only means whereby pressure can be directly measured in the glomerular capillaries and is thus essential in studies evaluating the contribution of the pre-and post-glomerular vessels to changes in renal hemodynamics and glomerular function 13,14.
To begin to investigate this hypothesis, we examined the effects of physiologically induced increases in renal sympathetic nerve activity (RSNA) in response to graded hypoxia on pre- and post-glomerular vascular resistances in anaesthetised rabbits10. We chose hypoxia to reflexly increase RSNA because we had previously shown that it produces graded increases in the amplitude of renal nerve firing (i.e. graded recruitment of individual nerves)44. Hypoxia has the further advantage that it does not significantly alter arterial pressure in the rabbit, thereby avoiding confounding autoregulatory effects on renal haemodynamics44. In the first study of its kind we measured simultaneously glomerular capillary pressure, renal nerve activity and whole kidney function, while subjecting the rabbits to different degrees of hypoxia10. The results were clear-cut, and compatible with the hypothesis that TYPE I and II axons can be differentially activated (see Fig. 3).
We found that moderate (14% O2) and severe (10% O2) hypoxia increased total RSNA by 60 % and 170 % respectively, chiefly by increasing the amplitude of the sympathetic bursts rather than their frequency. Moderate hypoxia decreased RBF (26%), increased glomerular capillary pressure and maintained GFR (Fig. 4). Both pre- and post-glomerular vascular resistances were increased; but there was a predominant effect on the post-glomerular vasculature. This greater effect on the efferent arteriole, when the TYPE II innervation density is similar on both afferent and efferent arterioles, can be explained on the basis of Poiseuille’s Law and the smaller resting diameter of the efferent arteriole10. In short, the recruitment of nerves in response to moderate hypoxia appeared to be predominantly TYPE II nerves (Fig. 3). In contrast, severe hypoxia decreased RBF (56%), with a significant fall in glomerular capillary pressure and GFR (Fig. 4). This pattern reflects a substantially greater pre-glomerular than post-glomerular vasoconstriction that is compatible with the further recruitment of nerves by severe hypoxia being predominantly TYPE I nerves (Fig. 3). These results provide evidence that different levels of reflexly induced increases in RSNA may differentially control pre- and post- glomerular vascular resistance, compatible with selective activation of TYPE I and II renal sympathetic nerves.
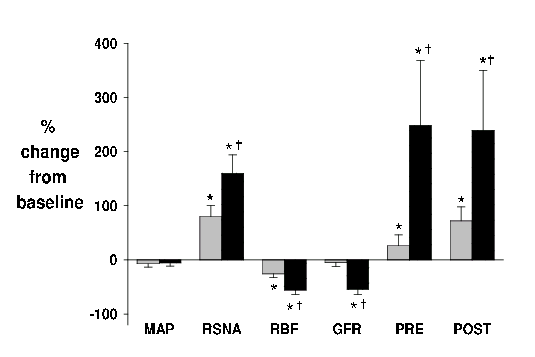
Figure 4. Responses to moderate (14% O2; grey) and severe (10% O2, black) hypoxia in anaesthetised rabbits. Values (means ± s.e.m. n = 7) are the percentage change from baseline (room air, 21% O2) for mean arterial pressure (MAP), renal sympathetic nerve activity (RSNA), renal blood flow (RBF), glomerular filtration rate (GFR) and pre (PRE) and post (POST glomerular vascular resistance. * P < 0.05 change from baseline, † P < 0.05 14% O2 vs 10% O2.
We are confident that the effects of hypoxia were mediated via the renal nerves as we have previously demonstrated the absence of any renal action of hypoxia following renal denervation42,45. However, neurally mediated renin release may have contributed to the response to increased RSNA, as renin cells are innervated by both TYPE I and II axons20. In particular, the renal response during moderate hypoxia might be explained on the basis of an increase in renin release being responsible for the rise in post-glomerular resistance. Though plasma renin activity was not increased in response to moderate hypoxia, intrarenal effects cannot be discounted10.
The question of the involvement of the renin-angiotensin system in the response to moderate (14% O2) hypoxia was investigated. The renin-angiotensin system was rendered unresponsive by the simultaneous infusion of an angiotensin converting enzyme inhibitor and ANGII to restore normal blood pressure (‘ANGII clamp’). Measurements were made in rabbits receiving either the ‘ANGII clamp’ or vehicle infusion before (room air, i.e. 21% O2) and during moderate hypoxia (14% O2) 46. As seen in our previous study10, in the vehicle group RSNA increased in response to 14% O2, and this decreased RBF, without effecting GFR or arterial pressure. Though the response was attenuated in the ‘ANGII clamp’ group, glomerular capillary pressure increased in both the vehicle and ‘ANGII clamp’ groups during 14% O2 (Fig. 5). These results are consistent with the notion that direct actions of TYPE II nerves on the efferent arterioles are responsible in part for the increase in post-glomerular resistance in response to 14% O2 46. These results further support our hypothesis that different populations of renal nerves selectively control pre- and post-glomerular resistance and hence glomerular pressure and ultrafiltration. Future studies will extend these findings by examining the renal microvascular response to stimulation of central nuclei involved in cardiovascular control31.
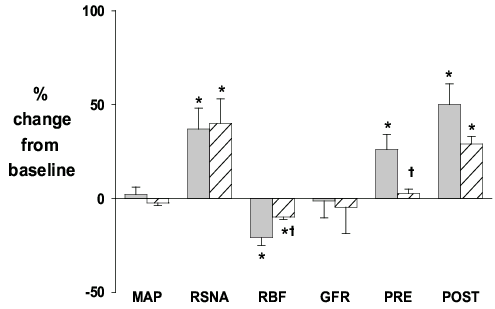
Figure 5. Responses to moderate (14% O2) hypoxia in anaesthetised rabbits treated throughout the study with vehicle (grey) or ‘ANGII clamp’ (infusion of an angiotensin converting enzyme inhibitor plus ANG II to restore blood pressure to normal; hatched). Values (means ± s.e.m. n = 6) are the percentage change from baseline (room air, 21% O2) for mean arterial pressure (MAP), renal sympathetic nerve activity (RSNA), renal blood flow (RBF), glomerular filtration rate (GFR) and pre (PRE) and post (POST glomerular vascular resistance. * P < 0.05 change from baseline, † P < 0.05 vehicle vs ANGII clamp.
Alterations in renal sympathetic nerve activity produce important effects on renal function, which contribute to the kidney’s main task of regulating body fluid balance. Our data suggest that there are functionally specific post-ganglionic renal nerves that can be selectively activated. Based on our evidence, we propose that TYPE II nerves predominate in the physiological control of arteriole resistance to maintain GFR constant during daily activity, whereas Type I nerves play a role when the animal is under stress (hemorrhage, dehydration or exercise), when blood flow is required for other organs at the expense of renal function. Overactivity of the renal nerves has been implicated in the pathophysiology of hypertension7, congestive heart failure6 and chronic renal failure8. Studies examining whether one or other of the populations of renal nerves are involved in these diseases offer possibilities of new therapeutic targets.
1. Bernard C. Lecon sur les proprietes physiologiques et les alterations pathologiques des liquides de l'organisme, Vol. 2. Bailliere et Fils, Paris. 1859; 170-171.
2. Smith HW. The kidney: Structure and function. Oxford University Press, New York, USA. 1951.
3. Anonymous. The wisdom of the kidney. N. Engl. J. Med. 1969; 280:106-107.
4. Barajas L, Muller J. The innervation of the juxtaglomerular apparatus and surrounding tubules: A quantitative analysis by serial section electron microscopy. J. Ultrastruct. Res. 1973; 43:107-132.
5. DiBona GF. Renal innervation and denervation: Lessons from renal transplantation reconsidered. Artif. Organs. 1987; 11:457-462.
6. DiBona G, Sawin L. Role of renal nerves in sodium retention of cirrhosis and congestive heart failure. Am. J. Physiol. 1991; 260:R298-R305.
7. Esler M. Sympathetic nervous system: Contribution to human hypertension and related cardiovascular diseases. J. Cardiovasc. Pharmacol. 1995; 26(suppl 2):S24-28.
8. Orth SR, Amann K, Strojek K, Ritz E. Sympathetic overactivity and arterial hypertension in renal failure. Nephrol. Dial. Transplant. 2001; 16:67-69.
9. Malpas S, Leonard BL. Neural regulation of renal blood flow; a re-examination. Clin. Exp. Pharmacol. Physiol. 2000; 27:956-964.
10. Denton KM, Shweta A, Anderson WP. Preglomerular and postglomerular resistance responses to different levels of sympathetic activation by hypoxia. J. Am. Soc. Nephrol. 2002; 13:27-34.
11. Maddox DA, Brenner BM. Glomerular ultrafiltration. In: Brenner BM (ed). Brenner & Rector's The Kidney, 6th edn, Vol 1. WB Saunders, Philedelphia. 2000; 286-333.
12. Kriz W, Gretz N, Lemley KV. Progression of glomerular diseases: Is the podocyte the culprit? Kid. Int. 1998; 54:687-697.
13. Arendshorst WJ, Gottschalk CW. Glomerular ultrafiltration dynamics: Historical perspective. Am. J. Physiol. 1985; 248:F163-174.
14. Denton KM, Anderson WP. Glomerular ultrafiltration in rabbits with superficial glomeruli. Pflügers. Arch. 1991; 419:235-242.
15. Trueta J, Barclay AE, Daniel PM, Franklin KJ, Pritchard MML. Studies of the renal circulation. Blackwell Scientific Publications. Oxford. 1947.
16. Kaissling B, Kriz W. Structural analysis of the rabbit kidney. Adv. Anat. Embryol. Cell. Biol. 1979; 56:7-12.
17. Denton KM, Anderson WP, Sinniah R. Effects of angiotensin II on regional afferent and efferent arteriole dimensions and the glomerular pole. Am. J. Physiol. 2000; 279:R629-638.
18. Denton KM, Fennessy PA, Alcorn D, Anderson WP. Morphometric analysis of the actions of angiotensin ii on renal arterioles and glomeruli. Am. J. Physiol. 1992; 262:F367-372.
19. Barajas L, Liu L, Powers K. Anatomy of the renal innervation: Intrarenal aspects and ganglia of origin. Can. J. Physiol. Pharmacol. 1992; 70:735-749.
20. Luff SE, Hengstberger SG, McLachlan EM, Anderson WP. Distribution of sympathetic neuroeffector junctions in the juxtaglomerular region of the rabbit kidney. J. Auton. Nerv. Syst. 1992; 40:239-254.
21. DiBona GF, Kopp UC. Neural control of renal function. Physiol. Rev. 1997; 77:75-197.
22. Luff SE. Ultrastructure of sympathetic axons and their structural relationship with vascular smooth muscle. Anat. Embryol. 1996; 193:515-531.
23. Luff SE, Hengstberger SG, McLachlan EM, Anderson WP. Two types of sympathetic axon innervating the juxtaglomerular arterioles of the rabbit and rat kidney differ structurally from those supplying other arteries. J. Neurocytol. 1991; 20:781-795.
24. DiBona GF, Sawin LL, Jones SY. Differentiated sympathetic neural control of the kidney. Am. J. Physiol. 1996; 271:R84-90.
25. Kopp UC, Dibona GF. Effects of the renal nerves and neurotransmitters on renal function. In: Brenner BM (ed). Brenner & Rector's The Kidney, 5th edn, Vol 1. WB Saunders, Philedelphia. 1996; 789-816.
26. Kopp UC, Dibona GF. The neural control of renal function. In: Seldin DW, Giebisch G, (eds). The Kidney, Physiology and Pathophysiology. 3rd edn. Vol. 1. Raven Press, New York. 2000; 1157-1204.
27. Grkovic I, Anderson CR. Calbindin D28k-immunoreactivity identifies distinct subpopulations of sympathetic pre- and postganglionic neurons in the rat. J. Comp. Neurol. 1997; 386:245-259.
28. Furness JB, Morris JL, Gibbins IL, Costa M. Chemical coding of neurons and plurichemical transmission. Annu. Rev. Pharmacol. Toxicol. 1989; 29:289-306.
29. Luff SE, Anderson WP, Denton KM, Young SB. Neuropeptide Y is specifically localised in type ii sympathetic axons innervating the afferent and efferent arterioles in the rabbit kidney. International Union of Physiological Sciences. 2001; ID1037 (Abstract).
30. Reinecke M, Forssmann WG. Neuropeptide (neuropeptide Y, neurotensin, vasoactive intestinal polypeptide, substance p, calcitonin gene-related peptide, somatostatin) immunohistochemistry and ultrastructure of renal nerves. Histochem. 1988; 89:1-9.
31. Dampney RA. Functional organization of central pathways regulating the cardiovascular system. Physiol. Rev. 1994; 74:323-364.
32. McAllen RM, Malpas SC. Sympathetic burst activity: Characteristics and significance. Clin. Exp. Pharmacol. Physiol. 1997; 24:791-799.
33. Kenney MJ, Weiss ML, Patel KP, Wang Y, Fels RJ. Paraventricular nucleus bicuculline alters frequency components of sympathetic nerve discharge bursts. Am. J. Physiol. 2001; 281:H1233-1241.
34. Dorward PK, Burke SL, Janig W, Cassell J. Reflex responses to baroreceptor, chemoreceptor, nociceptor inputs in single renal sympathetic neurones in the rabbit and the effects of anaesthesia on them. J. Auton. Nerv. System. 1987; 18:39-54.
35. Stein RD, Weaver LC. Multi- and single-fiber mesenteric and renal sympathetic responses to chemical stimulation of the intestinal receptors in cats. J. Physiol. 1988; 396:155-172.
36. Kon V. Neural control of renal circulation. Miner. Electrol. Metabol. 1989; 15:33-43.
37. Malpas SC, Ninomiya I. A new approach to analysis of synchronized sympathetic nerve activity. Am. J. Physiol. 1992; 263:H1311-1317.
38. Ninomiya I, Malpas SC, Matsukawa K, Shindo T, Akiyama T. The amplitude of synchronized cardiac sympathetic nerve activity reflects the number of activated pre- and postganglionic fibers in anesthetized cats. J. Auton. Nerv. System. 1993; 45:139-147.
39. Malpas SC, Evans RG. Do different levels and patterns of sympathetic activation all provoke renal vasoconstriction? J. Auton. Nerv. System. 1998; 69:72-82.
40. Handa RK, Johns EJ. A study of the renal responses in the rat to electrical stimulation of the afferent nerves of the brachial plexus. Quart. J. Exp. Physiol. 1988; 73:915-929.
41. Osborn JW, Livingstone RH, Schramm LP. Elevated renal nerve activity after spinal transection: Effects on renal function. Am. J. Physiol. 1987; 253:R619-R625.
42. Malpas SC, Shweta A, Anderson WP, Head GA. Functional response to graded increases in renal nerve activity during hypoxia in conscious rabbits. Am. J. Physiol. 1996; 271:R1489-1499.
43. Malpas SC, Head GA, Anderson WP. Renal responses to increases in renal sympathetic nerve activity induced by brainstem stimulation in rabbits. J. Auton. Nerv. System. 1996; 61:70-78.
44. Malpas SC, Bendle RD, Head GA, Ricketts JH. Frequency and amplitude of sympathetic discharges by baroreflexes during hypoxia in conscious rabbits. Am. J. Physiol. 1996; 271:H2563-2574.
45. Leonard BL, Malpas SC, Denton KM, Madden AC, Evans RG. Differential control of intrarenal blood flow during reflex increases in sympathetic nerve activity. Am. J. Physiol. 2001; 280:R62-68.
46. Denton KM, Flower RL, Anderson WP. Increased glomerular capillary pressure following moderate reflex activation of renal nerves is not due to angII; studies in rabbits with superficial glomeruli. Hypertension 2002; 40:408 (Abstract).