1. Influx of Ca2+, Mg2+ and Na+, and efflux of K+, have central importance for the function and survival of vascular smooth muscle cells, but progress in understanding the influx/efflux pathways has been restricted by a lack of identification of the genes underlying many of the non-voltage-gated cationic channels.
2. This review highlights evidence suggesting the genes are mammalian homologues of the Transient Receptor Potential (TRP) gene of the fruit-fly, Drosophila. The weight of evidence supports roles for TRPC1, TRPP2/1, and TRPC6, but recent studies point also to TRPC3, TRPC4/5, TRPV2, TRPM4, and TRPM7.
3. Activity of these TRP channels is suggested to modulate contraction and sense changes in intracellular Ca2+ storage, G-protein coupled receptor-activation, and osmotic stress. Roles in relation to myogenic tone, actions of vasoconstrictors substances, Mg2+-homeostasis, and the vascular injury response are suggested.
4. Knowledge that TRP channels are relevant to vascular smooth muscle cells in both their contractile and proliferative phenotypes should pave the way for a better understanding of vascular biology and provide the basis for discovery of a new set of therapeutic agents targeted to vascular disease.
Blood vessels are essential for the development and physiology of every organ of the body and vascular disease is the most common cause of death or disability in the western world1-5. Critical to all blood vessels except capillaries is the smooth muscle cell, a cell that is mostly restricted to its contractile phenotype in the adult but which undergoes profound phenotypic modulation in response to appropriate stimuli, providing a core element of vascular injury and adaptive responses, and underlying components of atherosclerotic plaques and neointimal hyperplasia.
Entry of the cations calcium (Ca2+), sodium (Na+) and magnesium (Mg2+) into the vascular smooth muscle cell is of central importance for the cell’s functional properties and phenotype6-13. Understanding these pathways has, however, been hampered by not knowing the identity of genes encoding numerous non-selective cationic channels - channels that are distinct from the voltage-gated Ca2+ channels, P2X receptors, and If pacemaker channels14-16. Existence of these additional cationic channels has been gradually appreciated over the past 30 years, largely because of the resistance of some signals to conventional Ca2+ antagonists such as verapamil, because electrophysiological studies show the signals are not voltage-activated, and because vasoconstrictor stimuli evoking the signals include not only ATP but also endothelin, noradrenaline, vasopressin, Ca2+-store depletion and membrane stretch15,17-19. A leap forward in understanding these pathways is now occurring following the discovery of TRP channels, the general properties of which have been reviewed20-25.
The TRP field originated in studies of Drosophila photoreceptors where a mutation in the TRP gene resulted in a more transient membrane potential depolarisation (transient receptor potential, TRP) in response to constant bright light, suggesting the mutation led to loss of function of an ion channel that normally mediates sustained depolarisation22. Since 1995, many mammalian homologues of the Drosophila TRP gene were reported and it became apparent that they, like Drosophila TRP, encode cationic channels20-25 – i.e. channels that confer membrane permeability to Na+ and K+, in many cases also Ca2+, and in at least one case, Mg2+. The mammalian TRP channels number at least 25 and are sub-classified based on amino acid sequence identity into groups denoted by an upper-case letter: For example, C for canonical or classical (hence TRPC1, TRPC2 … TRPC7), indicating the highest similarity to the archetypal Drosophila TRP; the full subclassification has been reviewed15,23-25. Some TRP channels are modulated by voltage, but depolarisation is not a critical trigger for channel opening, as it is, for example, in voltage-gated Ca2+ channels. Instead, intriguing activation signals have emerged, including temperature change, hydrogen peroxide, and gustatory chemicals. Although much attention has focused on the roles of mammalian TRP channels in sensory systems, the channels are also prevalent elsewhere and in vascular smooth muscle they are providing fertile ground for discovery. This review provides a summary of progress, highlighting evidence that at least ten types of TRP channel have functions in either the contractile or proliferative phenotype of the vascular smooth muscle cell – roles that are both physiological and potentially pivotal in vascular disease.
TRPC1 was the first mammalian homologue of Drosophila TRP to be cloned20,21 and throughout the subsequent decade TRPC1 was widely studied. Nevertheless, TRPC1 has, in some regards, been a frustrating subject because its heterologous over-expression has often not produced a signal clearly distinguishable from background signals (data have been reviewed26). Therefore, signals reported for TRPC1 may be obfuscated by endogenous ion transport mechanisms of the expression system, and it remains unclear if TRPC1 can actually function without the co-operation of other proteins, including other TRP channels. For example, it is plausible that TRPC1 operates only as part of a protein complex; for example, as per KV9.3 in complexes with KV2.1/2, and KIR6.1 in complexes with SUR227,28. Heterologous expression studies of TRPC1 have not produced agreement on the activation mechanism.
Much of the appreciation of TRPC1 comes from efforts to inhibit endogenously expressed TRPC1. These studies have been perhaps surprisingly successful, leading to agreement on the functional role of TRPC1 at the cellular level. Early on, mRNA encoding TRPC1 was found to be widely expressed, including in vascular smooth muscle15,26. Although there are fewer studies at the protein level it seems fair to say that TRPC1 protein is also widely expressed. Further, using antibody targeted to an extracellular epitope of TRPC1 we showed at least a fraction of TRPC1 in smooth muscle is a trans-plasma membrane protein mediating Ca2+-influx29. Tonic activity was not evident. Instead, activity occurred once Ca2+ stores were passively depleted using thapsigargin. Studies with antisense DNA and siRNA targeted to TRPC1 mRNA have yielded similar results for arterial smooth muscle cells growing in culture30,31. Therefore, three independent laboratories, using different techniques, conclude TRPC1 is a component of the store-operated channel (SOC) of the arterial smooth muscle cell in either its contractile or proliferative phenotype. There are no conflicting reports.
The broad structural similarity of TRPC1 to the Shaker voltage-gated K+ channel suggests TRPC1 is actually part of the ion permeation pathway of the SOC and not simply an accessory regulatory protein partner. This does not, however, exclude the involvement of other TRP proteins in the SOC, particularly because - again by analogy with Shaker-like channels - TRPC1 is likely to be only one component of a heterotetrameric channel complex. Heterologous expression studies and biochemical assays indicate TRPC1 can associate with TRPC4, TRPC5, TRPC3 and TRPP232-36. Indeed, siRNA targeted to mRNA encoding TRPC4 suppresses the SOC signal in pulmonary artery myocytes in culture, without effect on background Ca2+i or Ca2+-release signals37. An antisense DNA study on mesangial cells – cells with some relation to vascular smooth muscle – also supports a role for TRPC4 in SOCs38. TRPC5 may act similarly39 and has the capacity to respond to store-depletion when over-expressed in HEK-293 cells40.
It is often envisaged that the functional role of TRPC1 is as part of a mechanism to refill Ca2+-stores, and this might well be one of its roles. However, TRP channels have multiplicity of gating - also called versatility of gating40-42; that is, they can be activated by more than one stimulus, perhaps making them integrators or coincidence detectors (see below). Therefore, in line with the complex and often controversial data from heterologous expression studies, TRPC1 may have other activation mechanisms that do not require store-depletion. Indeed, endogenous SOCs in vascular smooth muscle are activated by noradrenaline independently of store-depletion43. The scope of TRPC1 activation mechanisms has probably yet to be discovered.
The overall importance of TRPC1 to vascular smooth muscle seems considerable – relating both to the muscle’s contractile and proliferative functions. Block of TRPC1 inhibits endothelin-evoked contraction in rat caudal artery44. Intriguingly, there is, in contrast, no role for TRPC1 in endothelin-evoked contraction of the basilar artery, unless it is subjected to organ culture, when store-operated Ca2+ entry in the smooth muscle cells is up-regulated45. In support of a role of TRPC1 in evoking contraction, over-expression of TRPC1 enhances pulmonary artery contraction evoked by cyclopiazonic acid (an agent that depletes Ca2+ stores) but not potassium-induced depolarisation46.
It is emerging that TRPC1 is not only relevant to Ca2+ entry that impacts on the contractile proteins. In part this suggestion has come from work on SOC in cerebral arterioles: TRPC1 is involved in this SOC29 but the SOC-mediated Ca2+-entry fails to evoke contraction, even though the Ca2+ elevation is equivalent to that evoked by 33 mM K+, which does cause contraction47. Although this Ca2+-entry might be tightly coupled to refilling of Ca2+ stores, as suggested by Casteels & Droogmans48, it may have other functions that are not directly related to generating contraction. Another function may be regulation of cell proliferation because antisense DNA targeted to TRPC1 mRNA inhibits proliferation of pulmonary artery smooth muscle cells in culture30. Furthermore, TRPC1 expression is up-regulated under conditions that evoke smooth muscle adaptation and proliferation30,31,45.
Polycystic kidney disease results from mutations in TRPP genes49. Intriguingly, these patients suffer not only from cysts on the kidney, but also from aneurysmal disease – thinning of the arterial wall that leads to rupture and internal bleeding. Similarly, mice with disrupted TRPP genes suffer from haemorrhage as well as cysts on the kidneys. The vascular effects relate in part to abnormality in vascular smooth muscle cells because TRPP1 and TRPP2 are expressed in these cells and SOC Ca2+ entry is suppressed in TRPP2+/- mice50-51. The link between this abnormality in Ca2+ handling and aneurysmal disease is unknown, but the suggestion is that TRPP proteins are involved in controlling the myo-elastic structural integrity of blood vessels52. Knowledge that TRPP2 binds to the C-terminus of TRPC132 indicates these proteins may have a co-operative relationship, but experimental data to test this idea are lacking.
It is widely agreed that TRPC6 is expressed in vascular smooth muscle15 and several studies have provided evidence of its functional importance. Strikingly, the functions proposed are diverse: Antisense DNA targeted to TRPC6 mRNA inhibits proliferation as well as remodelling of pulmonary artery smooth muscle cells53. It also inhibits store-operated Ca2+ entry in pulmonary myocytes53, cerebral artery myogenic tone54, and phenylephrine-evoked cationic current and Ca2+-entry in portal vein smooth muscle cells55. Welsh et al.54 also found cationic channel activation by hypo-osmotic stress was suppressed by antisense DNA to TRPC6. siRNA targeted to mRNA encoding TRPC6 also suppressed cation entry evoked by the diacylglycerol OAG31. Although not shown directly, vasopressin-evoked activation of cationic channels in A7r5 cells may also be mediated by TRPC656.
There is, therefore, the suggestion that endogenous TRPC6 is a store-operated, receptor-operated, stretch-activated and osmotically-activated cationic channel in vascular smooth muscle cells. Although multiplicity of activation is an emerging concept for TRP channels40,42 extensive studies of heterologously over-expressed TRPC6 show it to be primarily linked to receptor activation, but not store-depletion, membrane stretch, or osmotic stress. TRPC6 can, however, be activated by diacylglycerols (albeit at high concentration), the arachidonic acid metabolite 20-HETE, or phosphatidylinositol 3,4,5-trisphosphate15,57, any of which could be a common factor in endogenous activation by more general stimuli. In this context, the effect of 20-HETE is intriguing because it is a potent vasoconstrictor, acting in part by inhibiting the BKCa potassium channel in vascular smooth muscle cells58. Whether 20-HETE activates cationic channels in vascular smooth muscle cells, as it does in cultured tracheal myocytes59, is not reported. 20-HETE is produced during the development of myogenic tone, although Scotland et al.60 recently suggested its effect is mediated via TRPV1 in perivascular sensory neurones.
If there turns out to be no common activation signal mediating the functional modalities of endogenous TRPC6 or it truly lacks multiplicity of gating, how should we explain the published data? We perhaps should not exclude that the antisense DNA probes to TRPC6 mRNA could lack specificity, or the expression of TRPC6 gene may be directly linked to other genes, leading to knock-on effects on other pathways – note for example a report on siRNA knock-down of TRPM761. Recently, Lin et al.31 found no effect of siRNA targeted to TRPC6 on SOC in pulmonary artery smooth muscle cells grown in culture, contrasting with data produced with antisense DNA to TRPC6 in essentially the same cell type53. We should also consider that heterologous over-expression in HEK 293 cells may fail to reproduce the contextual environment of the smooth muscle cell – obviously HEK 293 cells do fail at this, but how important is this failure? It is clear that TRPC6 is highly relevant to smooth muscle function but we have much to understand about its regulation and roles.
Idiopathic pulmonary arterial hypertension (IPAH) is caused by excessive smooth muscle proliferation and results in right heart failure. Kunichika et al.62 and Yu et al.63 found IPAH to be associated with up-regulated expression of TRPC6 and TRPC3 (but not TRPC1) genes at the levels of mRNA and protein. These effects occur in the smooth muscle cells and siRNA targeted to mRNA encoding TRPC6 is an inhibitor of proliferation in smooth muscle cells cultured from patients with IPAH. Furthermore, chronic hypoxia enhances TRPC6 expression31 and the endothelin receptor antagonist bosentan, which is used in the treatment of IPAH, has a marked suppressive effect on TRPC6 expression62. Therefore, whatever the endogenous activation mechanism of TRPC6, there are indications of the channel’s importance in pulmonary vascular disease. A functional role of TRPC3 in these events has not been demonstrated, but it can form heteromultimers with TRPC636 and may have particular relevance because of its capacity to enable tonic entry of cations64.
TRPC3 has been commonly detected in smooth muscle15 but no functional roles had been assigned until recently. It is now apparent that antisense DNA targeted to mRNA encoding TRPC3 inhibits depolarisation and vasoconstriction evoked by uridine triphosphate (UTP), but not luminal pressure, in rat cerebral artery65. The antisense DNA also inhibited UTP-evoked whole cell current in isolated smooth muscle cells. Therefore, a role for TRPC3 in agonist evoked cationic current and contraction is suggested. Although TRPC3 can associate with TRPC6 and confer tonic cationic channel activity64, no role for TRPC3 in determination of the resting membrane potential or myogenic tone was evident in the studies of Reading et al.65. Further, although TRPC6 is receptor-operated (see above), and expressed in cerebral arteries, the UTP-evoked cationic current is abolished by antisense DNA targeted to TRPC3.
TRPV2 was first associated with sensory systems but its mRNA is detected in a wide range of other cell types66, including blood vessels67. In murine aortic myocytes, cell-swelling caused by hypotonic solution activates non-selective cationic channels and evokes Ca2+ entry67. These responses are inhibited by ruthenium red, a blocker of TRPV channels. Treatment of mouse aorta with antisense DNA targeted to TRPV2 mRNA reduced the amount of TRPV2 protein and suppressed activation of cationic channels by hypo-osmotic stimulation. Muraki et al.67 showed TRPV2 over-expressed in Chinese hamster ovary (CHO) cells is activated not only by hypo-osmotic stimulation but also by mechanical deformation of the platform on which the cells were grown. The implication of these findings for the general function of blood vessels remains to be elucidated. For example, is TRPV2 involved in generation of myogenic tone? Muraki et al.67 also detected mRNA encoding TRPV4 in aorta, although at lower levels than for TRPV2. The functional significance of this finding was not described, but it may be relevant that a TRPV4-like signal is detected in airways smooth muscle cells grown in culture68.
There have been reports of endogenous Ca2+-activated cationic channels in vascular smooth muscle69-71. The presence of such channels might serve to amplify vasoconstrictor signals and so the expression of TRP channels that are Ca2+-activated is of potential interest. Although TRPC5 is stimulated by intracellular calcium34,40, the most striking Ca2+-activated TRP channels would seem to be those in the TRPM subfamily. The first found to be strongly activated by intracellular Ca2+ was TRPM272,73 - expression of which occurs, at least at mRNA level, in arterial smooth muscle74. For TRPM2, the cofactor ADP-ribose is thought to be necessary for Ca2+-activation to manifest itself. TRPM4 can be activated by intracellular Ca2+ in the absence of a co-factor, although there is a rapid decline in Ca2+-sensitivity when membrane-patches are excised from cells, as if a co-factor is normally involved75. It has been proposed that TRPM8 is a ‘coincidence’ detector – i.e. that it senses intracellular Ca2+ if a co-factor (an icilin-like agent in the case of TRPM8) is present76. This might be a common theme for TRPM channels.
Messenger RNA encoding TRPM4 has been detected along with a Ca2+-activated cationic channel in smooth muscle cells of cerebral arteries71. High micromolar concentrations of intracellular Ca2+ are required to activate this channel, which may raise questions about the physiological relevance of the effect, or over whether Ca2+ is the normal activation signal, or whether a co-factor is required. The latter is indicated because it was observed that detection of the endogenous TRPM4-like channels was more reliable after prior treatment of cells with a phorbol ester.
Antisense DNA targeted to TRPM4 mRNA has been introduced into organ cultured arteries and caused near-abolition of myogenic tone, rather as antisense DNA to TRPC6 does in the same arteries54. Therefore although TRPM4 is not known to be stretch activated, it some how contributes critically to the myogenic response, perhaps working co-operatively with TRPC6 or in series with it. One hypothesis is that Ca2+ entering through TRPC6 channels stimulates TRPM4 channels such that TRPM4 is an amplification mechanism for TRPC6. TRPM4 is permeable to Na+ but not calcium77.
TRPM7 was originally noted for its importance in cell survival and its ubiquitous distribution78. Subsequently it was shown to confer membrane permeability to a wide range of cations and suggested to be an endogenous Mg2+ channel, regulating the intracellular Mg2+ concentration and consequently cell survival79,80. There is now evidence these concepts are directly applicable to vascular smooth muscle cells growing in culture, where TRPM7 gene is expressed as mRNA and protein81. siRNA targeted to mRNA encoding TRPM7 nearly abolished the acute elevation in the intracellular Mg2+ concentration when extracellular Mg2+ was elevated from 0 to 2 mM. Furthermore, angiotensin II and aldosterone enhanced TRPM7 gene expression, and the chronic positive effect of angiotensin II on intracellular Mg2+ levels was inhibited by the siRNA targeted to TRPM7. The siRNA also suppressed angiotensin II-evoked cell proliferation. TRPM7 would therefore seem to be a prime regulator of Mg2+ homeostasis in vascular smooth muscle, which is likely to have considerable importance given the wide-ranging vascular effects of magnesium12,82,83.
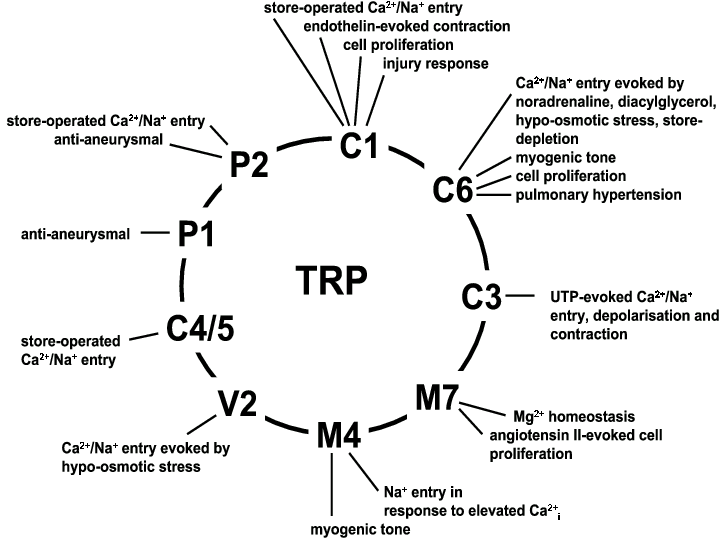
Figure 1 provides a summary of the emerging evidence suggesting that at least 10 of the 25, or so, TRP channels have functional roles in vascular smooth muscle. Such an extensive display of TRP function is impressive and challenges progress on any other cell type of the body. More research will of course be needed to confirm and further explore the roles of these, and other, TRP channels. In order to achieve this, development of more and better tools to explore the channels will be an on-going requirement – including the development of selective pharmacological agents. We also need a better understanding of the activation mechanisms of the channels, identification of endogenous regulatory ligands, and more efforts to appreciate the heteromultimerisation of endogenous TRP channels and their association with protein complexes. We are also challenged with understanding the overlapping functions and multifunctional nature of the TRP channels. Nevertheless, these are exciting and progressive times in this field. The knowledge that TRP channels are relevant to vascular smooth muscle function in both its contractile and proliferative phenotypes should pave the way for a better understanding of vascular biology and provide new targets for agents that might reduce the death and disability rates resulting from vascular disease.
Figure 1. Emerging functions of 10 types of TRP cationic channel in vascular smooth muscle. The TRP channels are shown pictorially arranged in a circle in recognition of their membership of one protein family. The circle does not necessarily indicate a physical association with an adjacent TRP. Nevertheless, there is experimental evidence for physical associations between TRPC1 and TRPP2, TRPC1, TRPC3, TRPC4 and TRPC5, and TRPC3 and TRPC6 (see text for references).
Supported by the Wellcome Trust, British Heart Foundation, and British and Australian Physiological Societies.
1. Balk EM, Karas RH, Jordan HS, Kupelnick B, Chew P, Lau J. Effects of statins on vascular structure and function: a systematic review. Am. J. Med. 2004; 117(10):775-790.
2. Dzau VJ, Braun-Dullaeus RC, Sedding DG. Vascular proliferation and atherosclerosis: new perspectives and therapeutic strategies. Nat. Med. 2002; 8(11):1249-1256.
3. D'Amore PA, Ng YS. Won't you be my neighbor? Local induction of arteriogenesis. Cell 2002; 110(3):289-292.
4. Risau W. Mechanisms of angiogenesis. Nature 1997; 386(6626):671-674.
5. Stocker R, Keaney JF, Jr. Role of oxidative modifications in atherosclerosis. Physiol. Rev. 2004; 84(4):1381-1478.
6. Kahl CR, Means AR. Regulation of cell cycle progression by calcium/calmodulin-dependent pathways. Endocr. Rev. 2003; 24(6):719-736.
7. Magnier-Gaubil C, Herbert JM, Quarck R, et al. Smooth muscle cell cycle and proliferation. Relationship between calcium influx and sarco-endoplasmic reticulum Ca2+ ATPase regulation. J. Biol. Chem. 1996; 271(44):27788-27794.
8. Munaron L. Calcium signalling and control of cell proliferation by tyrosine kinase receptors. Int. J. Mol. Med. 2002; 10(6):671-676.
9. Quignard JF, Ryckwaert F, Albat B, Nargeot J, Richard S. A novel tetrodotoxin-sensitive Na+ current in cultured human coronary myocytes. Circ. Res. 1997; 80(3):377-382.
10. Quignard JF, Harricane MC, Menard C, et al. Transient down-regulation of L-type Ca2+ channel and dystrophin expression after balloon injury in rat aortic cells. Cardiovasc. Res. 2001; 49(1):177-188.
11. Taurin S, Dulin NO, Pchejetski D, et al. c-Fos expression in ouabain-treated vascular smooth muscle cells from rat aorta: evidence for an intracellular-sodium-mediated, calcium-independent mechanism. J. Physiol. 2002; 543(Pt 3):835-847.
12. Touyz RM, Yao G. Modulation of vascular smooth muscle cell growth by magnesium-role of mitogen-activated protein kinases. J. Cell Physiol. 2003; 197(3):326-335.
13. Yoo HJ, Kozaki K, Akishita M, et al. Augmented Ca2+ influx is involved in the mechanism of enhanced proliferation of cultured vascular smooth muscle cells from spontaneously diabetic Goto-Kakizaki rats. Atherosclerosis 1997; 131(2):167-175.
14. Albert AP, Large WA. Store-operated Ca2+-permeable non-selective cation channels in smooth muscle cells. Cell Calcium 2003; 33(5-6):345-356.
15. Beech DJ, Muraki K, Flemming R. Non-selective cationic channels of smooth muscle and the mammalian homologues of Drosophila TRP. J. Physiol. 2004; 559(Pt 3):685-706.
16. Large WA. Receptor-operated Ca2+-permeable nonselective cation channels in vascular smooth muscle: a physiologic perspective. J. Cardiovasc. Electrophysiol. 2002; 13(5):493-501.
17. Bolton TB. Mechanisms of action of transmitters and other substances on smooth muscle. Physiol. Rev. 1979; 59(3):606-718.
18. McFadzean I, Gibson A. The developing relationship between receptor-operated and store-operated calcium channels in smooth muscle. Br. J. Pharmacol. 2002; 135(1):1-13.
19. Van Breemen C, Aaronson P, Loutzenhiser R. Sodium-calcium interactions in mammalian smooth muscle. Pharmacol. Rev. 1978; 30(2):167-208.
20. Zhu X, Chu PB, Peyton M, Birnbaumer L. Molecular cloning of a widely expressed human homologue for the Drosophila trp gene. FEBS. Lett. 1995; 373(3):193-198.
21. Wes PD, Chevesich J, Jeromin A, Rosenberg C, Stetten G, Montell C. TRPC1, a human homolog of a Drosophila store-operated channel. Proc. Natl. Acad. Sci. U S A. 1995; 92(21):9652-9656.
22. Minke B, Cook B. TRP channel proteins and signal transduction. Physiol. Rev. 2002; 82(2):429-472.
23. Montell C, Birnbaumer L, Flockerzi V. The TRP channels, a remarkably functional family. Cell 2002; 108(5):595-598.
24. Vennekens R, Voets T, Bindels RJ, Droogmans G, Nilius B. Current understanding of mammalian TRP homologues. Cell Calcium 2002; 31(6):253-264.
25. Clapham DE. TRP channels as cellular sensors. Nature 2003; 426(6966):517-524.
26. Beech DJ, Xu SZ, McHugh D, Flemming R. TRPC1 store-operated cationic channel subunit. Cell Calcium 2003; 33(5-6):433-440.
27. Babenko AP, Aguilar-Bryan L, Bryan J. A view of sur/KIR6.X, KATP channels. Annu. Rev. Physiol. 1998; 60:667-687.
28. Salinas M, Duprat F, Heurteaux C, Hugnot JP, Lazdunski M. New modulatory alpha subunits for mammalian Shab K+ channels. J. Biol. Chem. 1997; 272(39):24371-24379.
29. Xu SZ, Beech DJ. TrpC1 is a membrane-spanning subunit of store-operated Ca2+ channels in native vascular smooth muscle cells. Circ. Res. 2001; 88(1):84-87.
30. Sweeney M, Yu Y, Platoshyn O, Zhang S, McDaniel SS, Yuan JX. Inhibition of endogenous TRP1 decreases capacitative Ca2+ entry and attenuates pulmonary artery smooth muscle cell proliferation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002; 283(1):144-155.
31. Lin MJ, Leung GP, Zhang WM, et al. Chronic hypoxia-induced upregulation of store-operated and receptor-operated Ca2+ channels in pulmonary arterial smooth muscle cells: a novel mechanism of hypoxic pulmonary hypertension. Circ. Res. 2004; 95(5):496-505.
32. Tsiokas L, Arnould T, Zhu C, Kim E, Walz G, Sukhatme VP. Specific association of the gene product of PKD2 with the TRPC1 channel. Proc. Natl. Acad. Sci. U S A. 1999; 96(7):3934-3939.
33. Lintschinger B, Balzer-Geldsetzer M, Baskaran T, et al. Coassembly of Trp1 and Trp3 proteins generates diacylglycerol- and Ca2+-sensitive cation channels. J. Biol. Chem. 2000; 275(36):27799-27805.
34. Strubing C, Krapivinsky G, Krapivinsky L, Clapham DE. TRPC1 and TRPC5 form a novel cation channel in mammalian brain. Neuron 2001; 29(3):645-655.
35. Strubing C, Krapivinsky G, Krapivinsky L, Clapham DE. Formation of novel TRPC channels by complex subunit interactions in embryonic brain. J. Biol. Chem. 2003; 278(40):39014-39019.
36. Hofmann T, Schaefer M, Schultz G, Gudermann T. Subunit composition of mammalian transient receptor potential channels in living cells. Proc. Natl. Acad. Sci. U S A. 2002; 99(11):7461-7466.
37. Zhang S, Remillard CV, Fantozzi I, Yuan JX. ATP-induced mitogenesis is mediated by cyclic AMP response element-binding protein-enhanced TRPC4 expression and activity in human pulmonary artery smooth muscle cells. Am. J. Physiol. Cell Physiol. 2004; 287(5):C1192-1201.
38. Wang X, Pluznick JL, Wei P, Padanilam BJ, Sansom SC. TRPC4 forms store-operated Ca2+ channels in mouse mesangial cells. Am. J. Physiol. Cell Physiol. 2004; 287(2):C357-364.
39. Beech DJ, Xu SZ, McHugh D, Zeng F, Mair L, Sivaprasadarao A. Antibody to the predicted outer pore of TRPC5 ablates calcium entry evoked by store-depletion in isolated rabbit arterioles. J. Physiol. 2004; 557:C78.
40. Zeng F, Xu SZ, Jackson PK, et al. Human TRPC5 channel activated by a multiplicity of signals in a single cell. J. Physiol. 2004; 559(Pt 3):739-750.
41. Lievremont JP, Bird GS, Putney JW, Jr. Canonical transient receptor potential TRPC7 can function as both a receptor- and store-operated channel in HEK-293 cells. Am. J. Physiol. Cell Physiol. 2004; 287(6):C1709-1716.
42. Vriens J, Watanabe H, Janssens A, Droogmans G, Voets T, Nilius B. Cell swelling, heat, and chemical agonists use distinct pathways for the activation of the cation channel TRPV4. Proc. Natl. Acad. Sci. U S A. 2004; 101(1):396-401.
43. Albert AP, Large WA. Activation of store-operated channels by noradrenaline via protein kinase C in rabbit portal vein myocytes. J. Physiol. 2002; 544(Pt 1):113-125.
44. Bergdahl A, Gomez MF, Dreja K, et al. Cholesterol depletion impairs vascular reactivity to endothelin-1 by reducing store-operated Ca2+ entry dependent on TRPC1. Circ. Res. 2003; 93(9):839-847.
45. Bergdahl A, Gomez MF, Wihlborg AK, et al. Plasticity of TRPC expression in arterial smooth muscle: correlation with store-operated Ca2+ entry. Am. J. Physiol. Cell Physiol. 2004 (In Press, published on-line).
46. Kunichika N, Yu Y, Remillard CV, Platoshyn O, Zhang S, Yuan JX. Overexpression of TRPC1 enhances pulmonary vasoconstriction induced by capacitative Ca2+ entry. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004; 287(5):962-969.
47. Flemming R, Cheong A, Dedman AM, Beech DJ. Discrete store-operated calcium influx into an intracellular compartment in rabbit arteriolar smooth muscle. J. Physiol. 2002; 543(Pt 2):455-464.
48. Casteels R, Droogmans G. Exchange characteristics of the noradrenaline-sensitive calcium store in vascular smooth muscle cells or rabbit ear artery. J. Physiol. 1981; 317:263-279.
49. Cantiello HF. Regulation of calcium signaling by polycystin-2. Am. J. Physiol. Renal Physiol. 2004; 286(6):F1012-1029.
50. Qian Q, Hunter LW, Li M, et al. Pkd2 haploinsufficiency alters intracellular calcium regulation in vascular smooth muscle cells. Hum. Mol. Genet. 2003; 12(15):1875-1880.
51. Qian Q, Li M, Cai Y, et al. Analysis of the polycystins in aortic vascular smooth muscle cells. J. Am. Soc. Nephrol. 2003; 14(9):2280-2287.
52. Kim K, Drummond I, Ibraghimov-Beskrovnaya O, Klinger K, Arnaout MA. Polycystin 1 is required for the structural integrity of blood vessels. Proc. Natl. Acad. Sci. U S A. 2000; 97(4):1731-1736.
53. Yu Y, Sweeney M, Zhang S, et al. PDGF stimulates pulmonary vascular smooth muscle cell proliferation by upregulating TRPC6 expression. Am. J. Physiol. Cell. Physiol. 2003; 284(2):C316-330.
54. Welsh DG, Morielli AD, Nelson MT, Brayden JE. Transient receptor potential channels regulate myogenic tone of resistance arteries. Circ. Res. 2002; 90(3):248-250.
55. Inoue R, Okada T, Onoue H, et al. The transient receptor potential protein homologue TRP6 is the essential component of vascular α(1)-adrenoceptor-activated Ca2+-permeable cation channel. Circ. Res. 2001; 88(3):325-332.
56. Jung S, Strotmann R, Schultz G, Plant TD. TRPC6 is a candidate channel involved in receptor-stimulated cation currents in A7r5 smooth muscle cells. Am. J. Physiol. Cell. Physiol. 2002; 282(2):C347-359.
57. Tseng PH, Lin HP, Hu H, Wang C, Zhu MX, Chen CS. The canonical transient receptor potential 6 channel as a putative phosphatidylinositol 3,4,5-trisphosphate-sensitive calcium entry system. Biochemistry 2004; 43(37):11701-11708.
58. Ma YH, Gebremedhin D, Schwartzman ML, et al. 20-Hydroxyeicosatetraenoic acid is an endogenous vasoconstrictor of canine renal arcuate arteries. Circ. Res. 1993; 72(1):126-136.
59. Cloutier M, Campbell S, Basora N, Proteau S, Payet MD, Rousseau E. 20-HETE inotropic effects involve the activation of a nonselective cationic current in airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003; 285(3):560-568.
60. Scotland RS, Chauhan S, Davis C, et al. Vanilloid receptor TRPV1, sensory C-fibers, and vascular autoregulation: a novel mechanism involved in myogenic constriction. Circ. Res. 2004; 95(10):1027-1034.
61. Aarts M, Iihara K, Wei WL, et al. A key role for TRPM7 channels in anoxic neuronal death. Cell 2003; 115(7):863-877.
62. Kunichika N, Landsberg JW, Yu Y, et al. Bosentan inhibits transient receptor potential channel expression in pulmonary vascular myocytes. Am. J. Respir. Crit. Care Med. 2004; 170(10):1101-1107.
63. Yu Y, Fantozzi I, Remillard CV, et al. Enhanced expression of transient receptor potential channels in idiopathic pulmonary arterial hypertension. Proc. Natl. Acad. Sci. U S A. 2004; 101(38):13861-13866.
64. Dietrich A, Mederos y Schnitzler M, Emmel J, Kalwa H, Hofmann T, Gudermann T. N-linked protein glycosylation is a major determinant for basal TRPC3 and TRPC6 channel activity. J. Biol. Chem. 2003; 278(48):47842-47852.
65. Reading SA, Earley S, Waldron BJ, Welsh DG, Brayden JE. TRPC3 Mediates Pyrimidine Receptor-induced Depolarization of Cerebral Artery. Am. J. Physiol. Heart Circ. Physiol. 2004. (In Press, published on-line)
66. O'Neil RG, Brown RC. The vanilloid receptor family of calcium-permeable channels: molecular integrators of microenvironmental stimuli. News Physiol. Sci. 2003; 18:226-231.
67. Muraki K, Iwata Y, Katanosaka Y, et al. TRPV2 is a component of osmotically sensitive cation channels in murine aortic myocytes. Circ. Res. 2003; 93(9):829-838.
68. Jia Y, Wang X, Varty L, et al. Functional TRPV4 channels are expressed in human airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004; 287(2):272-278.
69. Loirand G, Pacaud P, Baron A, Mironneau C, Mironneau J. Large conductance calcium-activated non-selective cation channel in smooth muscle cells isolated from rat portal vein. J. Physiol. 1991; 437:461-475.
70. Wang Q, Hogg RC, Large WA. A monovalent ion-selective cation current activated by noradrenaline in smooth muscle cells of rabbit ear artery. Pflugers Arch. 1993; 423(1-2):28-33.
71. Earley S, Waldron BJ, Brayden JE. Critical role for transient receptor potential channel TRPM4 in myogenic constriction of cerebral arteries. Circ. Res. 2004; 95(9):922-929.
72. Perraud AL, Fleig A, Dunn CA, et al. ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nature 2001; 411(6837):595-599.
73. McHugh D, Flemming R, Xu SZ, Perraud AL, Beech DJ. Critical intracellular Ca2+ dependence of transient receptor potential melastatin 2 (TRPM2) cation channel activation. J. Biol. Chem. 2003; 278(13):11002-11006.
74. Xu SZ, Beech DJ. Multiple trp genes expressed as mRNA and protein in rabbit and human arteries. J. Physiol. 2001; 535:S69.
75. Nilius B, Prenen J, Tang J, et al. Regulation of the Ca2+ sensitivity of the non-selective cation channel TRPM4. J. Biol. Chem. 2004. (In Press, published on-line)
76. Chuang HH, Neuhausser WM, Julius D. The super-cooling agent icilin reveals a mechanism of coincidence detection by a temperature-sensitive TRP channel. Neuron 2004; 43(6):859-869.
77. Launay P, Fleig A, Perraud AL, Scharenberg AM, Penner R, Kinet JP. TRPM4 is a Ca2+-activated nonselective cation channel mediating cell membrane depolarization. Cell 2002; 109(3):397-407.
78. Nadler MJ, Hermosura MC, Inabe K, et al. LTRPC7 is a Mg.ATP-regulated divalent cation channel required for cell viability. Nature 2001; 411(6837):590-595.
79. Monteilh-Zoller MK, Hermosura MC, Nadler MJ, Scharenberg AM, Penner R, Fleig A. TRPM7 provides an ion channel mechanism for cellular entry of trace metal ions. J. Gen. Physiol. 2003; 121(1):49-60.
80. Schmitz C, Perraud AL, Johnson CO, et al. Regulation of vertebrate cellular Mg2+ homeostasis by TRPM7. Cell 2003; 114(2):191-200.
81. He Y, Yao G, Savoia C, Touyz RM. Transient Receptor Potential Melastatin 7 Ion Channels Regulate Magnesium Homeostasis in Vascular Smooth Muscle Cells. Role of Angiotensin II. Circ. Res. 2004. (In Press, published on-line)
82. Altura BM, Altura BT. Magnesium and cardiovascular biology: an important link between cardiovascular risk factors and atherogenesis. Cell. Mol. Biol. Res. 1995; 41(5):347-359.
83. McHugh D, Beech DJ. Modulation of Ca2+ channel activity by ATP metabolism and internal Mg2+ in guinea-pig basilar artery smooth muscle cells. J. Physiol. 1996; 492 ( Pt 2):359-376.