1. β1-adrenergic and M2 muscarinic receptor regulation of cAMP production plays a pivotal role in autonomic regulation of cardiac myocyte function. However, not all responses are easily explained by a uniform increase or decrease in cAMP activity throughout the entire cell.
2. Adenovirus expression of fluorescence resonance energy transfer (FRET) based biosensors can be used to monitor cAMP activity in protein kinase A (PKA) signaling domains as well as the bulk cytoplasmic domain of intact adult cardiac myocytes.
3. Data obtained using FRET-based biosensors expressed in different cellular microdomains has been used to develop a computational model of compartmentalized cAMP signaling.
4. A systems biology approach that employs quantitative computational modeling together with experimental data obtained using FRET-based biosensors has been used to provide evidence for the idea that compartmentation of cAMP signaling is necessary to explain the stimulatory responses to β1-adrenergic receptor activation as well as the complex temporal responses to M2 muscarinic receptor activation.
Many different neurotransmitters and hormones are capable of regulating the electrical, mechanical, and metabolic activity of the heart by stimulating the production of cAMP. The signaling pathway typically involves receptor activation of adenylyl cyclase (AC) via a stimulatory G protein (Gs)-dependent mechanism. The receptor most commonly associated with activation of this signaling pathway in cardiac myocytes is the β1-adrenergic receptor (β1AR). However, not all receptors capable of stimulating cAMP production produce the same functional responses. The frequent explanation for such observations has been that different receptors are capable of stimulating cAMP production in distinct microdomains within the cell.1 The classic example of this is the fact that both β1ARs as well as E2/4 prostaglandin receptors are capable of stimulating cAMP production, but only β1AR activation leads to changes in acute functional responses.2-6
Still other types of receptors can inhibit as well as stimulate cAMP production, eliciting complex temporal responses. An example of this is the M2 muscarinic receptor (M2R).7-16 In ventricular myocytes, M2R activation alone has little or no effect on cell function. However, M2R activation can potently inhibit the stimulatory responses associated with β1AR activation of cAMP production by inhibiting AC activity via an inhibitory G protein (Gi)-dependent mechanism. This is referred to as accentuated antagonism.17 Furthermore, in the presence of submaximal β1AR activation, transient activation of M2Rs actually produces a biphasic effect.7-13 In the presence of muscarinic agonists such as acetylcholine (ACh), there is antagonism of the β-adrenergic response, but upon withdrawal of ACh, there is an exaggerated stimulatory or rebound response. This reflects the fact M2R activation of a Gi-dependent signaling pathway produces an inhibitory effect that turns on and off rapidly, as well as a stimulatory response that turns on and off more slowly. This complex temporal response to ACh is also difficult to explain if one assumes that receptor activation produces a uniform increase or decrease in cAMP throughout the cell.
Until recently it has only been possible to measure cAMP activity using biochemical methods that typically involve homogenization of tissue or cell preparations.1 While the importance of information obtained using such an approach cannot be over emphasized, obtaining evidence for compartmentation of cAMP signaling was somewhat limited by this approach. Homogenized preparations could be separated into soluble (cytosolic) and particulate (membrane) fractions by centrifugation. When this was done, β-adrenergic agonists were found to increase cAMP production and activation of protein kinase A (PKA) in both fractions, whereas prostaglandins only affected cAMP production and PKA activation in the soluble fraction.2,6,18,19 This was consistent with the observation that most functional responses were associated with cAMP-dependent activation of type II PKA, which is found primarily in the particulate fraction.1,20,21 These kinds of results seemed consistent with the idea that the difference in the responses to β-adrenergic agonists and prostaglandins could be explained by compartmentation of cAMP production. However, understanding how this related to what was happening in an intact cell was an open question.
More recently, several different approaches for measuring cAMP in intact cells have been developed. One class of cAMP biosensors utilizes the principle of fluorescence resonance energy transfer (FRET).22-24 One of the first FRET-based biosensors for measuring cAMP activity was constructed using PKA.25 In this case, a donor fluorophore, rhodamine, was covalently attached to the regulatory subunit of PKA and an acceptor fluorophore, fluorescein, was attached to the catalytic subunit. This probe was introduced into cells by microinjection or dialysis via a patch pipette.25-27 In the resting state, the two fluorophores are in close proximity (<100 Å) to one another. Under these conditions, excitation of the donor leads to direct transfer of energy to the acceptor resulting in its fluorescence. When cAMP levels increased, binding of the nucleotide to the regulatory subunit results in reorientation of the catalytic and regulatory subunits and a loss of FRET. By measuring the change in the donor/acceptor fluorescence ratio, it is possible to obtain a readout of changes in cAMP activity in an intact cell. More recently, genetically encoded versions of this type of biosensor have been developed,28,29 including one in which cyan (CFP) and yellow fluorescent protein (YFP) were used as the donor and acceptor fluorophores, respectively. Such probes have made it possible to introduce the biosensor in a variety of cell types using standard transfection techniques. By using the type II regulatory subunit of PKA, which contains peptide sequences that bind to A kinase anchoring proteins (AKAPs), this type of probe can then be expressed in the same pattern as endogenous type II PKA.28,30 As a result, this sensor is expected to respond to cAMP specifically in type II PKA signaling domains.
Other FRET based cAMP biosensors have been developed using the exchange protein activated by cAMP (Epac).31-33 The principle involved is the same as that for the PKA-based probe. An increase in cAMP activity results in a loss of FRET that can be used as an indicator of changes in cAMP activity. However, with the Epac-based probes, CFP and YFP are attached to the amino and carboxy termini of the same protein. There are various versions of Epac-based biosensors, but one created by Nikolaev et al.31 was constructed using only the cAMP binding domain of type 2 Epac. This Epac2 cAMP sensor (Epac2-camps) lacks any anchoring sequences that might target it to specific locations within the cell. The result is a biosensor that is able to diffuse freely throughout the cytoplasm.
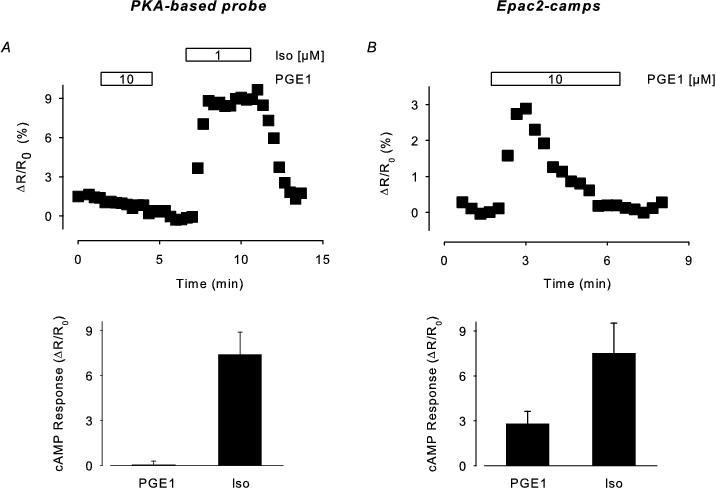
Figure 1. A. PKA-based biosensor expressed in adult guinea pig ventricular myocytes. Top panel: time course of changes in the CFP/YFP fluorescence ratio (ΔR) relative to baseline (R0) observed during exposure to PGE1 and isoproterenol (Iso). Bottom panel: average response to 10 μM PGE1 and subsequent exposure to 1 μM Iso (n=5). B. Epac2-camps expressed in adult guinea pig ventricular myocytes. Top panel: time course of changes in ΔR/R0 observed during exposure to 10 μM PGE1. Bottom panel: average response produced by 10 μM PGE1 (n=8) or 1 μM Iso (n=8). Data adapted from Warrier et al.34
In our initial studies, we tested the proof of concept that because the PKA-based probe and Epac2-camps exhibit different expression patterns, when expressed in adult ventricular myocytes, they would be able to differentiate between responses to different agonists. Type II PKA is found primarily in the particulate fraction of homogenized cardiac preparations. Therefore, we predicted that the type II PKA-based biosensor would detect responses to agonists that stimulate cAMP production in the particulate fraction of cells but not the soluble fraction. On the other hand, if the Epac2-based probe is freely diffusible, we predicted that it would respond to agonists that stimulate cAMP production in the soluble fraction.
We expressed these probes in adult guinea pig ventricular myocytes using an adenovirus based approach.34 In myocytes expressing the PKA-based probe, exposure to the prostaglandin receptor agonist PGE1 failed to elicit a detectable change in FRET, even though subsequent exposure to the β-adrenergic agonist isoproterenol (Iso) resulted in a significant response (Figure 1A). However, PGE1 was clearly able to stimulate cAMP production, because exposure of myocytes expressing the Epac2-based probe to PGE1 resulted in a significant, albeit transient response (Figure 1B). These results support the conclusion that FRET based biosensors expressed in different microdomains can be used to detect compartmentalized responses in intact cardiac myocytes. These results are also consistent with the idea that the PKA-based biosensor responds specifically to cAMP in a particulate or membrane domain associated with type II PKA that is not accessible to the soluble or bulk cytoplasmic domain through free diffusion.
The stimulatory and inhibitory effects of M2R activation can be explained in part by the different types of AC that are expressed in cardiac muscle and how they are regulated by M2R activation of Gi.8 Cardiac myocytes express AC types 5 and 6 (AC5/6) as well as AC types 4 and 7 (AC4/7).35 However, while the activated α subunit of Gi (Giα) inhibits AC5/6, it has no affect on AC4/7.36-38 In fact, AC4/7 activity is actually stimulated by Gi βγ subunits.39,40 In this way, activation of the M2R can both inhibit as well as stimulate cAMP production (see Figure 2). However, this still does not readily explain the difference in the speed of the stimulatory and inhibitory responses.
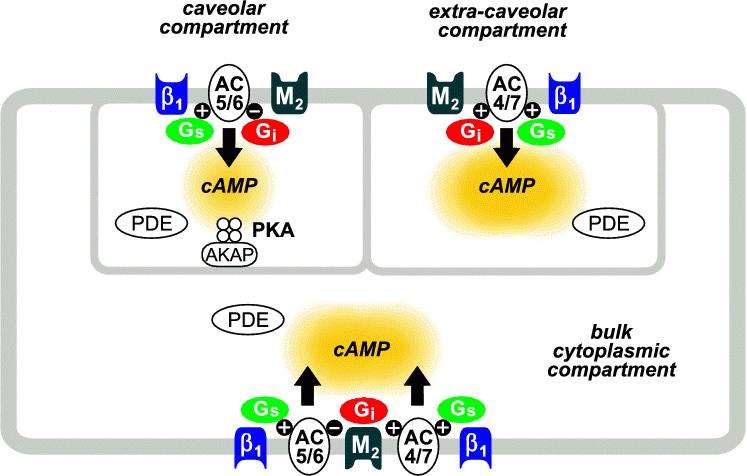
Figure 2. Model of compartmentalized cAMP signaling pathways in a cardiac ventricular myocyte. β1-adrenergic receptor (β1), M2 muscarinic receptor (M2), stimulatory (Gs) and inhibitory (Gi) G-proteins, adenylyl cyclase types 5 and 6 (AC5/6), adenylyl cyclase types 4 and 7 (AC4/7), phosphodiesterase (PDE).
A clue as to the potential explanation for the complex temporal response has come from studies demonstrating that AC5/6 is expressed in cholesterol rich fractions of the plasma membrane specifically associated with caveolin-3, a scaffolding protein that is involved in forming caveolae.41-43 On the other hand, AC4/7 appears to be found primarily in cholesterol rich lipid rafts that do not include caveolin-3.43,44 This suggests that M2R inhibition and stimulation of cAMP may be occurring in different subcellular locations. Inhibition of cAMP production occurs in a caveolar domain. This is also where type II PKA is found,42,45 and again, activation of type II PKA correlates closely with regulation of functional responses.1,20,21 Stimulation of cAMP production occurs in an extracaveolar membrane compartment. Based on this, we hypothesized that the rebound stimulatory effect that is associated with functional responses is due to the time-dependent flux of cAMP from an extracaveolar to a caveolar compartment.
In order to test our hypothesis for the complex response to M2R activation, we developed a computational model of compartmentalized cAMP signaling in a cardiac myocyte.46 This model was created using the wealth of quantitative kinetic data existing in the literature. The compartmentalized nature of the model was made possible because of more recent information describing the relative distribution of the various proteins involved in cAMP signaling between cytosolic and membrane fractions as well as among different membrane fractions of cardiac myocytes. The key element in this case is the inclusion of AC5/6 activity in the caveolar domain and AC4/7 activity in the extracaveolar domain. The model assumes that there are three compartments (Figure 2). The first compartment is a caveolar domain that includes 10% of the plasma membrane and makes up 1% of the total cell volume. The second compartment is an extracaveolar domain that includes 20% of the plasma membrane and makes up 2% of the cell volume. The third compartment is the bulk cytoplasmic domain that is associated with the remainder of the plasma membrane and makes up 97% of the cell volume.
We then took advantage of results obtained using the PKA-based biosensor to validate the model. This probe appears to selectively respond to cAMP in a type II PKA signaling domain (see Figure 1). Furthermore, type II PKA is associated with the membrane fraction of cardiac myocytes in general and the caveolar membrane fraction in particular.42,45 Therefore, we compared the responses detected by the PKA-based probe to those predicted in the caveolar domain of the model. We were able to demonstrate that the model is able to accurately describe the concentration dependence of responses to β1AR activation. It is also able to reproduce both the inhibitory and stimulatory responses produced by M2R activation in the presence of β1AR stimulation.46
One advantage of using a computational approach is that it makes it possible to see what is happening in different microdomains in order to get a better understanding of the model’s ability to produce different behaviours. If we look specifically at AC5/6 and AC4/7 activity as well as cAMP levels in the caveolar and extracaveolar domains (Figure 3), we can see that exposure to a submaximally stimulating concentration of Iso results in a slight increase in cAMP production by all AC isoforms, which is associated with a slight increase in cAMP levels in both the caveolar and extracaveolar domains. However, subsequent exposure to ACh results in an inhibition of cAMP production by AC5/6 and cAMP levels in the caveolar domain, while there is a significant stimulation of cAMP production by AC4/7 and cAMP levels in the extracaveolar domain. This creates a significant gradient in cAMP concentrations, which results in a flux of cAMP from the extracaveolar to caveolar compartments. Although this causes a slight increase in cAMP in the caveolar domain, the actual concentration of cAMP is assumed to be below the threshold for producing functional responses. Upon withdrawal of ACh and termination of M2R activation, there is a rapid reversal of AC5/6 inhibition as well as AC4/7 stimulation. However, the cAMP gradient does not dissipate immediately. The result is that extracaveolar cAMP continues to diffuse into the caveolar domain, resulting in cAMP levels transiently increasing to a level much higher than that observed prior to M2R activation. Eventually, phosphodiesterase (PDE) activity breaks down the excess cAMP, explaining the transient nature of the stimulatory response.
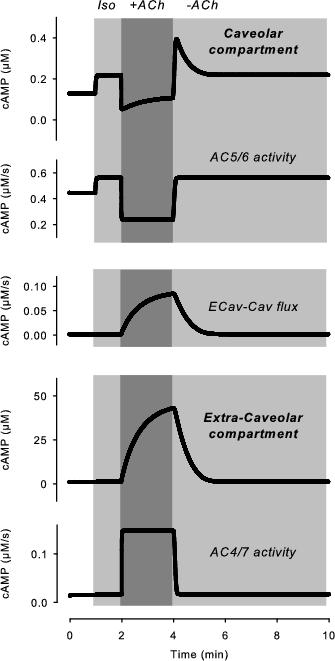
Figure 3. Kinetics of changes in cAMP activity associated with muscarinic stimulatory response. Changes in caveolar and extracaveolar cAMP production and concentration caused by transient exposure to acetylcholine (ACh) in the presence of a submaximally stimulating concentration of isoproterenol (Iso). Adapted from Iancu et al.46
The development of cAMP biosensors provides a means for directly measuring cAMP activity in different subcellular locations within live cells. This can be helpful in demonstrating that different receptors regulate cAMP responses in distinct microdomains. These biosensors have also been helpful in developing a quantitative computational approach to understanding more complex behaviours.
One outcome of this type of systems biology approach is the generation of non-intuitive predictions that can then be tested experimentally. For example, results obtained from the original version of our model suggest that the basal level of cAMP in the bulk cytoplasmic compartment of cardiac ventricular myocytes should be ∼1 μM. This is significantly higher than the ∼100 nM levels suggested to exist in the caveolar domain by measurements obtained using the PKA-based biosensor.46 This implication is important because it suggests that compartmentation plays an important role even under basal conditions by maintaining microdomains where cAMP levels are significantly below that found throughout most of the cell. This allows receptor signaling to regulate cAMP concentration over a range that modulates the activity of high affinity effectors such as type II PKA, which has a Kd of 300 nM.47
A second unexpected prediction of the model has to do with the role of PDE activity in creating the microdomains involved in compartmentation of β1AR and M2R responses. It is often suggested that PDEs act as functional barriers responsible for creating compartments.1,48 Consistent with this idea, different PDE isoforms have been shown to be targeted to specific subcellular locations.47,49 Furthermore, the non-uniform distribution of PDE activity between compartments of our model is essential for determining the concentration of cAMP within each compartment, and therefore the gradients between compartments. However, PDE activity alone is not sufficient to maintain those gradients.46 The model predicts that there must be some other factor contributing to the limited diffusion of cAMP between these compartments.
It is important to note that our computational model is a working hypothesis. It can provide a theoretical framework for testing the feasibility of complex hypotheses. Yet, the model itself is only as good as the data on which it is based. Therefore, it is imperative to continually evaluate the model using new experimental data. In this respect, the use of different cAMP biosensors should only improve our ability to generate more quantitative means of studying cell signaling. This should also facilitate our ability to expand the model to include other signaling pathways that regulate cAMP activity. For example, the current version of the model only contains the β1 subtype of β-adrenergic receptor. This allows direct comparison of the model’s output with experimental results obtained using guinea pig ventricular myocytes, which only express this subtype of βAR.50,51 However, cardiac myocytes from most mammals also express a significant subpopulation (10-20%) of β2ARs.1 Furthermore, β2ARs are able to produce compartmentalized cAMP-dependent responses because of their ability to couple to Gi as well as Gs signaling pathways.1,52 Therefore, a systems biology approach to studying cAMP signaling may provide a useful means of investigating the diversity of response produced by many different types of receptors.
1. Steinberg SF, Brunton LL. Compartmentation of G protein-coupled signaling pathways in cardiac myocytes. Annu. Rev. Pharmacol. Toxicol. 2001; 41: 751-773.
2. Hayes JS, Brunton LL, Brown JH, Reese JB, Mayer SE. Hormonally specific expression of cardiac protein kinase activity. Proc. Natl. Acad. Sci. USA. 1979; 76: 1570-1574.
3. Brunton LL, Hayes JS, Mayer SE. Hormonally specific phosphorylation of cardiac troponin I and activation of glycogen phosphorylase. Nature 1979; 280: 78-80.
4. Keely SL. Activation of cAMP-dependent protein kinase without a corresponding increase in phosphorylase activity. Res. Commun. Chem. Pathol. Pharmacol. 1977; 18: 283-290.
5. Brunton LL, Hayes JS, Mayer SE. Functional compartmentation of cyclic AMP and protein kinase in heart. Adv. Cyclic Nucleotide Res. 1981; 14: 391-397.
6. Hayes JS, Brunton LL, Mayer SE. Selective activation of particulate cAMP-dependent protein kinase by isoproterenol and prostaglandin E1. J. Biol. Chem. 1980; 255: 5113-5119.
7. Zakharov SI, Harvey RD. Rebound stimulation of the cAMP-regulated Cl- current by acetylcholine in guinea-pig ventricular myocytes. J. Physiol. (Lond.) 1997; 499: 105-120.
8. Belevych AE, Sims C, Harvey RD. ACh-induced rebound stimulation of L-type Ca2+ current in guinea-pig ventricular myocytes, mediated by Gβγ-dependent activation of adenylyl cyclase. J. Physiol. (Lond.) 2001; 536: 677-692.
9. Belevych AE, Harvey RD. Muscarinic inhibitory and stimulatory regulation of the L-type Ca2+ current is not altered in cardiac ventricular myocytes from mice lacking endothelial nitric oxide synthase. J. Physiol. (Lond.) 2000; 528: 279-289.
10. Song Y, Shryock JC, Belardinelli L. Potentiating effect of acetylcholine on stimulation by isoproterenol of L-type Ca2+ current and arrhythmogenic triggered activity in guinea pig ventricular myocytes. J. Cardiovasc. Electrophysiol. 1998; 9: 718-726.
11. Ehara T, Mitsuiye T. Adrenergic-cholinergic interactions on membrane potential of K+-depolarized ventricular muscle. Am. J. Physiol. 1984; 247: H244-H250.
12. Tareen FM, Ono K, Noma A, Ehara T. β-adrenergic and muscarinic regulation of the chloride current in guinea-pig ventricular cells. J. Physiol. (Lond.) 1991; 440: 225-241.
13. Ono K, Noma A. Autonomic regulation of cardiac chloride current. Jpn. J. Physiol. 1994; 44 Suppl 2: S193-8.
14. Hollenberg M, Carriere S, Barger AB. Biphasic action of acetylcholine on ventricular myocardium. Circ. Res. 1965; 16: 527-536.
15. Endoh M. Muscarinic regulation of Ca2+ signaling in mammalian atrial and ventricular myocardium. Eur. J. Pharmacol. 1999; 375: 177-196.
16. Gilmour RF, Zipes DP. Positive inotropic effect of acetylcholine in canine cardiac Purkinje fibers. Am. J. Physiol. 1985; 249: H735-H740.
17. Harvey RD, Belevych AE. Muscarinic regulation of cardiac ion channels. Br. J. Pharmacol. 2003; 139: 1074-1084.
18. Hayes JS, Brunton LL. Functional compartments in cyclic nucleotide action. J. Cyclic Nucleotide Res. 1982; 8: 1-16.
19. Buxton IL, Brunton LL. Compartments of cyclic AMP and protein kinase in mammalian cardiomyocytes. J. Biol. Chem. 1983; 258: 10233-10239.
20. Aass H, Skomedal T, Osnes JB. Increase of cyclic AMP in subcellular fractions of rat heart muscle after β-adrenergic stimulation: prenalterol and isoprenaline caused different distribution of bound cyclic AMP. J. Mol. Cell. Cardiol. 1988; 20: 847-860.
21. Hohl CM, Li Q. Compartmentation of cAMP in adult canine ventricular myocytes: Relation to single-cell free Ca2+ transients. Circ. Res. 1991; 69: 1369-1379.
22. Zaccolo M. Use of chimeric fluorescent proteins and fluorescence resonance energy transfer to monitor cellular responses. Circ. Res. 2004; 94: 866-873.
23. Nikolaev VO, Lohse MJ. Monitoring of cAMP synthesis and degradation in living cells. Physiology (Bethesda) 2006; 21: 86-92.
24. Nikolaev VO, Bunemann M, Schmitteckert E, Lohse MJ, Engelhardt S. Cyclic AMP imaging in adult cardiac myocytes reveals far-reaching β1-adrenergic but locally confined β2-adrenergic receptor-mediated signaling. Circ. Res. 2006; 99: 1084-1091.
25. Adams SR, Harootunian AT, Buechler YJ, Taylor SS, Tsien RY. Fluorescence ratio imaging of cyclic AMP in single cells. Nature 1991; 349: 694-697.
26. Bacskai BJ, Hochner B, Mahaut-Smith M, Adams SR, Kaang BK, Kandel ER, Tsien RY. Spatially resolved dynamics of cAMP and protein kinase A subunits in Aplysia sensory neurons. Science 1993; 260: 222-226.
27. Goaillard JM, Vincent PV, Fischmeister R. Simultaneous measurements of intracellular cAMP and L-type Ca2+ current in single frog ventricular myocytes. J. Physiol. (Lond.) 2001; 530: 79-91.
28. Zaccolo M, Pozzan T. Discrete microdomains with high concentration of cAMP in stimulated rat neonatal cardiac myocytes. Science 2002; 295: 1711-1715.
29. Zaccolo M, De Giorgi F, Cho CY, Feng L, Knapp T, Negulescu PA, Taylor SS, Tsien RY, Pozzan T. A genetically encoded, fluorescent indicator for cyclic AMP in living cells. Nat. Cell Biol. 2000; 2: 25-29.
30. Warrier S, Belevych AE, Ruse M, Eckert RL, Zaccolo M, Pozzan T, Harvey RD. β-adrenergic and muscarinic receptor induced changes in cAMP activity in adult cardiac myocytes detected using a FRET based biosensor. Am. J. Physiol. 2005; 289: C455-C461.
31. Nikolaev VO, Bunemann M, Hein L, Hannawacker A, Lohse MJ. Novel single chain cAMP sensors for receptor-induced signal propagation. J. Biol. Chem. 2004; 279: 37215-37218.
32. DiPilato LM, Cheng X, Zhang J. Fluorescent indicators of cAMP and Epac activation reveal differential dynamics of cAMP signaling within discrete subcellular compartments. Proc. Natl. Acad. Sci. USA. 2004; 101: 16513-16518.
33. Ponsioen B, Zhao J, Riedl J, Zwartkruis F, van der KG, Zaccolo M, Moolenaar WH, Bos JL, Jalink K. Detecting cAMP-induced Epac activation by fluorescence resonance energy transfer: Epac as a novel cAMP indicator. EMBO Rep. 2004; 5: 1176-1180.
34. Warrier S, Ramamurthy G, Eckert RL, Nikolaev VO, Lohse MJ, Harvey RD. cAMP microdomains and L-type Ca2+ channel regulation in guinea-pig ventricular myocytes. J. Physiol. 2007; 580: 765-776.
35. Ishikawa Y, Homcy CJ. The adenylyl cyclases as integrators of transmembrane signal transduction. Circ. Res. 1997; 80: 297-304.
36. Sunahara RK, Dessauer CW, Gilman AG. Complexity and diversity of mammalian adenylyl cyclases. Annu. Rev. Pharmacol. Toxicol. 1996; 36: 461-480.
37. Taussig R, Gilman AG. Mammalian membrane-bound adenylyl cyclases. J. Biol. Chem. 1995; 270: 1-4.
38. Tang WJ, Gilman AG. Adenylyl cyclases. Cell 1992; 70: 869-872.
39. Chen J, DeVivo M, Dingus J, Harry A, Li J, Sui J, Carty DJ, Blank JL, Exton JH, Stoffel RH, Inglese J, Lefkowitz RJ, Logothetis DE, Hildebrandt JD, Iyengar R. A region of adenylyl cyclase 2 critical for regulation by G protein beta gamma subunits. Science 1995; 268: 1166-1169.
40. Federman AD, Conklin BR, Schrader KA, Reed RR, Bourne HR. Hormonal stimulation of adenylyl cyclase through Gi-protein β γ subunits. Nature 1992; 356: 159-161.
41. Head BP, Patel HH, Roth DM, Lai NC, Niesman IR, Farquhar MG, Insel PA. G-protein-coupled receptor signaling components localize in both sarcolemmal and intracellular caveolin-3-associated microdomains in adult cardiac myocytes. J. Biol. Chem. 2005; 280: 31036-31044.
42. Rybin VO, Xu X, Lisanti MP, Steinberg SF. Differential targeting of β-adrenergic receptor subtypes and adenylyl cyclase to cardiomyocyte caveolae. A mechanism to functionally regulate the cAMP signaling pathway. J. Biol. Chem. 2000; 275: 41447-41457.
43. Ostrom RS, Insel PA. The evolving role of lipid rafts and caveolae in G protein-coupled receptor signaling: implications for molecular pharmacology. Br. J. Pharmacol. 2004; 143: 235-245.
44. Cooper DM. Regulation and organization of adenylyl cyclases and cAMP. Biochem. J. 2003; 375: 517-529.
45. Balijepalli RC, Foell JD, Hall DD, Hell JW, Kamp TJ. Localization of cardiac L-type Ca2+ channels to a caveolar macromolecular signaling complex is required for β2-adrenergic regulation. Proc. Natl. Acad. Sci. USA. 2006; 103: 7500-7505.
46. Iancu RV, Jones SW, Harvey RD. Compartmentation of cAMP signaling in cardiac myocytes: a computational study. Biophys J. 2007; 92: 3317-3331.
47. Mongillo M, McSorley T, Evellin S, Sood A, Lissandron V, Terrin A, Huston E, Hannawacker A, Lohse MJ, Pozzan T, Houslay MD, Zaccolo M. Fluorescence resonance energy transfer-based analysis of cAMP dynamics in live neonatal rat cardiac myocytes reveals distinct functions of compartmentalized phosphodiesterases. Circ. Res. 2004; 95: 67-75.
48. Brunton LL. PDE4: arrested at the border. Sci. STKE. 2003; 2003: E44.
49. Fischmeister R, Castro LR, bi-Gerges A, Rochais F, Jurevicius J, Leroy J, Vandecasteele G. Compartmentation of cyclic nucleotide signaling in the heart: the role of cyclic nucleotide phosphodiesterases. Circ. Res. 2006; 99: 816-828.
50. Hool LC, Harvey RD. Role of β1- and β2-adrenergic receptors in regulation of Cl- and Ca2+ channels in guinea pig ventricular myocytes. Am. J. Physiol. 1997; 273: H1669-H1676.
51. Ranu HK, Mak JC, Barnes PJ, Harding SE. Gi-dependent suppression of β1-adrenoceptor effects in ventricular myocytes from NE-treated guinea pigs. Am. J. Physiol. Heart Circ. Physiol. 2000; 278: H1807-H1814.
52. Xiao RP. β-adrenergic signaling in the heart: dual coupling of the β2-adrenergic receptor to Gs and Gi proteins. Sci STKE. 2001; 2001: RE15.