When an actively contracting skeletal muscle is stretched, the force it exerts increases throughout the stretch due to the strain of contractile and non-contractile proteins (Edman, Elzinga & Noble, 1978). After the stretch this force enhancement decays to a new steady-state force, which is higher than the isometric force at the corresponding length, often referred to as residual force enhancement (RFE). The mechanism of RFE is still unknown, although previous studies suggest it may be caused by the strain of activated non-contractile proteins, such as titin (Pinniger, Ranatunga & Offer, 2006). In order to evaluate the possible role of titin in stretch-induced force enhancement we compared the force enhancement characteristics in fast (EDL) and slow (SOL) muscle, known to differ in their titin isoforms (Wang et al., 1991).
EDL and SOL muscles were removed from the hindlimb of adult male Wistar rats under deep anesthesia (IP sodium pentobarbitone, 40 mg/kg) and mounted in an in vitro muscle test system (1200A; Aurora Scientific, Aurora, Canada) containing mammalian Ringer solutions bubbled with 90% O2 / 5% CO2 at 25 °C. We examined the stretch-induced force response by applying a series of small amplitude stretches (5% optimal fibre length, L0) to the plateau of an isometric contraction at a range of constant velocities (0.1-10 L0.s-1). Based on the force-velocity profile we quantified the velocity-dependent contractile and velocity-independent non-contractile contributions to force enhancement during stretch, and RFE after stretch in EDL and SOL muscles (Pinniger, Ranatunga & Offer, 2006).
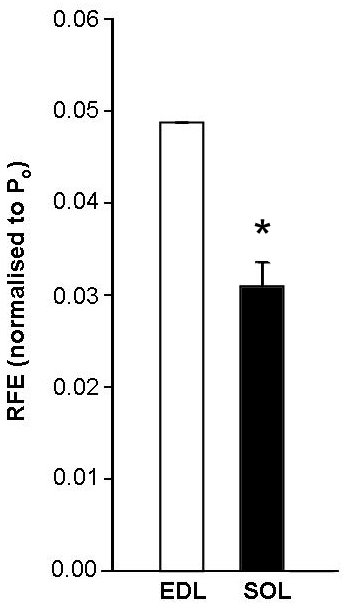
RFE was significantly higher in EDL than SOL (p < 0.01; see figure). This supports our hypothesis that the non-contractile contribution to force enhancement is higher in fast muscle and maybe due to a shorter, stiffer titin filament. We also examined the non-contractile contributions to stretch-induced force enhancement in damaged and dystrophic muscle. Following a series of 10 damaging eccentric contractions (20% L0) RFE increased significantly (3-4 times) in both EDL and SOL (n = 9, p < 0.01 ), which suggests that non-contractile proteins may be overstretched as a result of muscle damage. Furthermore, RFE was significantly lower in dystrophic muscles from mdx mice compared to non-dystrophic control (C57BL/10) mice in EDL (mdx 0.03 ± 0.005 Po (n = 4), control 0.06 ± 0.004 Po (n = 4), p < 0.01) and SOL (mdx (n = 4) 0.02 ± 0.002 Po, control 0.03 ± 0.003 Po (n = 4, p < 0.01) muscles.
The results of this study are consistent with the hypothesis that titin plays a significant role in stretch-induced force enhancement. Titin is ideally situated within the sarcomere to act as a sensor for mechanical strain that may, via complex interactions with other sarcomeric proteins, be crucial for the activation of various signalling pathways associated with muscle development and adaptation. Based on the changes observed in the magnitude of RFE we propose that the activation of these pathways may be enhanced after muscle damage and impaired in diseased muscle, e.g., Duchenne muscular dystrophy.
Edman K., Elzinga G & Noble M. (1978) Journal of Physiology, 281: 139-55.
Pinniger G, Ranatunga K & Offer G. (2006) Journal of Physiology 573: 627-43.
Wang K, McCarter R, Wright J, Beverly J & Ramirez-Mitchell R. (1991) Proceedings of the National Academy of Sciences USA, 88: 7101-5.