1. Aquaporins are targets for drug discovery for basic research and medicine. Human diseases involving fluid imbalances and oedema are of major concern, and involve tissues in which aquaporins are expressed. The range of functional properties of aquaporins is continuing to expand steadily with ongoing research in the field.
2. Gating domains in aquaporins (AQPs) are molecular sites for drug actions. Discovery of the arylsulfonamide AqB013 as an antagonist for AQP1 and AQP4 provided the first pharmacological agent with translational promise for the treatment of diseases in which AQPs have been implicated. The putative binding site for AqB013 in the internal vestibule of the AQP water pore involves amino acid residues that are located in the AQP loop D gating domain.
3. Aquaporins have been proposed as novel targets in cancer and oedema, and are associated with a surprising array of important processes in the brain and body such as angiogenesis, cell migration, development, and neuropathological diseases. Functions beyond their simple role as water channels are suggested by the subtype-specific regulation of AQP expression. In both cancer and brain oedema, current therapies are limited, and new pharmacological approaches focused on AQPs offer exciting potential for clinical advances.
Aquaporins are integral membrane proteins that enable the movement of water and other small solutes across biological membranes.1 With continuing research on aquaporins, the repertoire now includes permeabilities in some subtypes of aquaporins to a variety of substrates including gasses2 and ions.3 As members of the Major Intrinsic Protein (MIP) family, aquaporins have been identified in almost every living organism, including vertebrates, invertebrates, plants, and bacteria. The 13 mammalian classes of aquaporins (AQP0-12) generally are divided into two categories: the aquaporins, which primarily transport water, and the aquaglyceroporins, which also allow permeation of small solutes such as glycerol and urea.
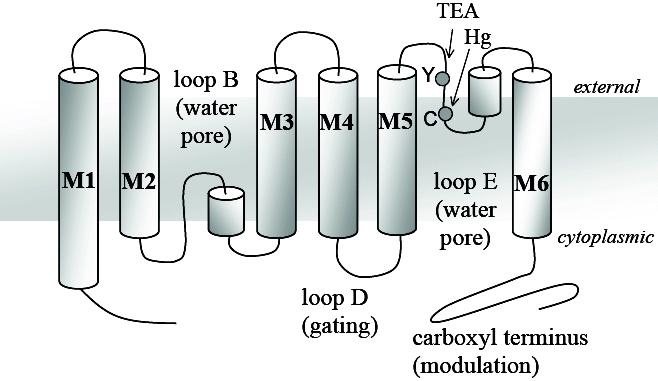
AQP1 was the first water channel to be characterized4,5 from studies of erythrocyte membrane proteins. Previously known as CHIP28 (channel-like integral protein, 28 kDa), Aquaporin-1 mediates osmotic and hydrostatic water fluxes across cell membranes. AQP1 is found in a variety of tissues including kidney proximal tubule, eye, heart, lung, the vascular system, and in the choroid plexus in brain ventricles.6-10 Exemplifying a general pattern of AQP structural organisation, each subunit in the AQP1 tetrameric channel comprises six full transmembrane domains, connected by intracellular and extracellular loops with cytoplasmic N- and C-termini (Figure 1). Domains within loop B (between transmembrane domains 2 and 3) and loop E (between transmembrane domains 5 and 6) fold into the lipid bilayer from opposite sides of the membrane to form the subunit aqueous pore, first conceptualized as an “hourglass” model11 as an insight that was generally validated by later crystal structural analyses.12,13
Figure 1. Transmembrane topology diagram of Aquaporin 1. This illustrates the six transmembrane domains (M1 to M6); the loops that form the intrasubunit water pore (loops B and E); and the regions implicated in gating (loop D), modulation of activity (carboxyl terminus), and block by the extracellular agents mercury (Hg) and tetraethylammonium ion (TEA).
Aquaporin blockers are expected to be valuable for intervention in diseases involving defects in fluid homeostasis.14,15 While the need for pharmacological tools to dissect the roles of aquaporins in physiological and pathological processes is clear, progress in the development of selective non-toxic pharmacological agents for AQP channels has been slow. The classic blocker of AQP1 is the mercuric ion Hg2+ 16 but its high toxicity precludes any therapeutic consideration. Site-directed mutagenesis of human AQP1 identified the binding site for Hg2+ as a cysteine residue (Cys189) located extracellularly in loop E,16 at which covalent modification is thought to block water permeability by occluding the pore. Other heavy metals such as silver and gold17 also inhibit AQP1 water channel function. The carbonic anhydrase inhibitor acetazolamide (10 μM) was found to reduce water permeability by >80% in AQP1-expressing Xenopus oocytes18 and in AQP1-transfected HEK cells.19
Prior work from our laboratory identified the quaternary ammonium, tetraethylammonium ion (TEA+), as the first non-mercurial compound to inhibit AQP1 function.20 With human AQP1 channels expressed in Xenopus oocytes, the application of extracellular TEA resulted in a modest dose-dependent reduction in osmotically-driven swelling rate, blocking water channel function by approximately 30% at 10 mM. TEA-Cl reduced water permeability in AQP1-expressing MDCK cells and in rat kidney descending thin limb21 indicating the block also was effective on natively expressed channels. The sensitivity of human AQP1 to TEA was removed by site-directed mutagenesis of Tyr186 to Phe (Y186F) in the outer vestibule of the AQP1 water pore, confirming that the inhibitory effect of TEA was mediated directly at the AQP1 channel.20 The block of AQP1 by TEA was confirmed independently, and extended to include other classes of AQPs which have tyrosine at the position homologous to Y186 in AQP1.22 Although a separate study failed to confirm TEA blocking activity,23 TEA has successfully been used as an aquaporin inhibitor in other preparations.24,25
The agents summarized above (metal ions, tetraethylammonium, acetazolamide) are valuable in that they indicate pharmacological modification of AQP activity is possible, but they lack the specificity or efficacy needed for a clear translational value. Our discovery of a novel blocker (AqB013) for AQP1 (Figure 2) and AQP4 channels, and the identification of the putative binding site on the intracellular vestibule of the water channel pore,26 suggest opportunities for translational applications. Targets of treatment include oedema in brain, lung and heart, glaucoma, cancer, renal dysfunction and other conditions involving alterations in fluid transport and homeostasis in AQP-expressing tissues.14
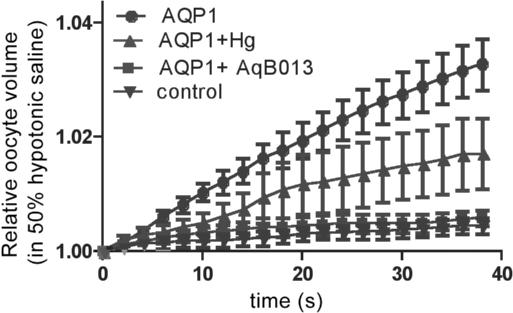
Figure 2. A novel blocker of mammalian aquaporin channels, AqB013, is more effective than mercury at the same dose (50 μM), as illustrated here for mammalian AQP1 channels in the Xenopus oocyte expression system. Oocytes were placed into 50% hypotonic saline at time zero. Data are mean ± SD, for 10 oocytes per treatment, with swelling rates calculated from the change in cross-sectional area as a function of time using video imaging. Control oocytes lack AQP channel expression and show minimal swelling in hypotonic conditions.
In the mammalian brain, the predominant class of water channel is AQP4, which is localized in astroglial cells throughout the brain and spinal cord near the blood-brain-barrier interface,27 and thought to play a major role in brain fluid homeostasis. It is also expressed in other tissues including lung, skeletal muscle and collecting ducts of the kidney.28,29 In contrast to AQP1, AQP4 is insensitive to mercurial inhibition.28,30 Atypically, rat AQP4 channels reconstituted in proteoliposomes show a mercury-sensitive water permeability that involves Cys178 in the loop D region,31 a site that normally is inaccessible on the intracellular side. The residue in the equivalent position in AQP1 is threonine (T157 in human AQP1).
Arylsulfonamides are a class of compounds characterized by an aromatic benzene ring attached to a sulfonamide group, SO2NH2 (Figure 3). This class of compounds shows promise for development of pharmacological agents for aquaporins. Sulfonamides such as the carbonic anhydrase inhibitor acetazolamide can reduce AQP4 activity.32 Anti-epileptic drugs such as topiramate and zonisamide recently have been found to have inhibitory effects on AQP4 function.33
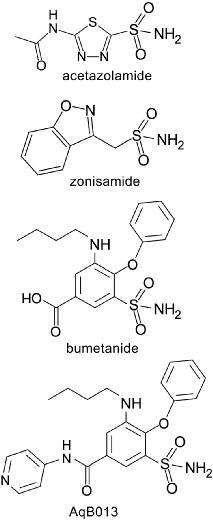
Figure 3. Chemical structures of arylsulfonamide compounds with aquaporin blocking activities.
Discovery of the arylsulfonamide AqB013 as an antagonist for AQP1 and AQP4 was based on a combination of synthetic chemistry, biological assays, and in silico modeling. Bumetanide was selected as the structural scaffold for design of a chemical library of derivatives based on a small but significant block of AQP4 water channel function.26 Bumetanide is known as a loop diuretic drug which inhibits the Na-K-2Cl cotransporter in the kidney ascending limb of Henle.34 Inhibiting the Na-K-2Cl cotransporter in the kidney decreases Na+ reabsorption and increases water excretion, resulting in diuresis. At doses higher than those needed for loop diuretic activity, bumetanide has a protective effect on rat brain oedema.35,36 Brain oedema and infarct volumes were reduced by application of approximately 100 μM bumetanide in cerebrospinal fluid saline,37,38 whereas the concentration required to block the cotransporter is more than 50-fold lower.34
The AqB series of bumetanide derivatives was tested on AQP1 and -4 channels expressed in Xenopus oocytes, assayed by videomicroscopic analysis of swelling rates. AqB013 produced half maximal block (IC50) at ∼20 μM for both AQP1 and -4, and had comparatively little effect on Na-K-2Cl transport. Collaborative studies on the effects of AqB013 in vivo and in vitro are in progress to assess effects of this compound in cell and systems models of pathophysiology. This novel class of pharmacological agents for aquaporins could be valuable as adjuncts in treating oedema and other conditions involving fluid imbalance in aquaporin-expressing tissues.
Crystal-structure data for AQPs allowed structural modelling and the prediction of putative binding sites on AQP4.26 In silico analyses were used to resolve three putative binding site cavities: two on the intracellular side of the AQP4 subunit, labelled as Sites 1 and 2, and one extracellular, labelled as Site 3 (Figure 4). Site-directed mutations of AQP4 were used to test for amino acid residues that contributed to the candidate binding sites. Changes at Site 1 altered efficacy of block by AqB013, thus showing that the aquaporin channel is the direct molecular target of action.
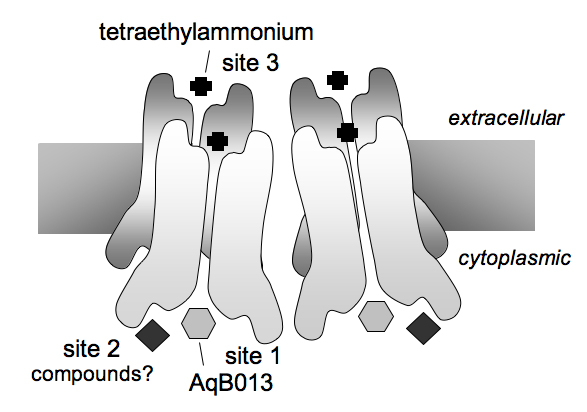
Figure 4. Schematic diagram of putative binding sites on the aquaporin subunit. Site 1 is the likely site of action of the novel aquaporin blocker AqB013, which is proposed to inhibit AQP water channel activity by occlusion of the internal vestibule. Site 2 also on the intracellular side of the AQP subunit is a candidate for possible modulatory compounds yet to be identified. In the extracellular vestibule, Site 3 is the likely site of action of quaternary ammonium compounds such as tetraethylammonium ion, as well as extracellular mercurial compounds.
Site 1 is located at the intracellular vestibule of the water pore, and involves the loop D region. The mutations of Ser180 to Arg (S180R), and Val189 to Ala (V189A) both significantly enhanced the blocking effect of 50 μM AqB013 as compared with wild type AQP4.26 The residues Ser180 and Val189 in rat AQP4 which interact with AqB013 are equivalent to Ser180 and Ile189 in human AQP4 (Figure 5), and are located in the intracellular loop D. AqB013 block at the equivalent position in human AQP1 would involve the loop D residues Arg159 and Ala168, which were recapitulated in the selection of substitute residues for the AQP4 modifications. The wild type AQP1 channel with the Arg and Ala residues naturally in place does not show higher sensitivity to block by AqB013 as compared with AQP4, suggesting that additional residues contribute to the putative docking site.
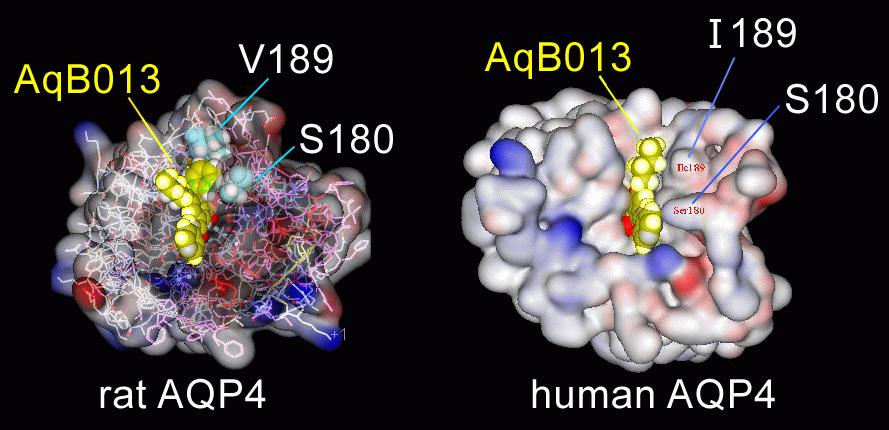
Figure 5. Structural models of rat and human AQP4. The view is from the cytoplasmic side of the subunits, showing the simulated docking of AqB013 in the internal vestibule of the water pore (Site 1), and the positions of two key residues (Ser180 and Val/Ile189) that were demonstrated by mutagenesis to influence the efficacy of the blocking effect.26
The loop D domain has been proposed to serve in gating AQP channel functions in mammalian AQP1,39 and AQP4,40,41 and in plant AQP.42 In AQP1, the loop D domain is involved in regulation of a non-selective monovalent cation channel activity, activated by cGMP signalling. Although AQP1 is a robust water channel, under permissive conditions, it also functions as an ion channel.9,43-45 The subunits of the aquaporin channel are organized as tetramers spanning the cell membrane, and the passage of water and ions appears to be through pharmacologically distinct pathways, with conduction of ions occurring through the putative central pore sensitive to Cd2+ but not TEA+ 39,46 and water transport occurring through the individual pores located in each subunit sensitive to TEA+ but not Cd2+.11 Development of therapeutic strategies could involve differential targeting of the AQP1 dual ion-and-water channel functions.
The candidate residues of Sites 1 and 2 are located mainly in intracellular loops B and D, but Site 2 also includes the C-terminal region of AQP4, based on structural modelling. The C-terminal region has been proposed to be involved in AQP1 regulation because it carries a consensus sequence for Ca2+ binding.41 As illustrated by the effects of site-directed mutations, the C-terminal domain modulates the ionic conductance of the AQP1 channel.39,45,47,48 Activation of protein kinase C by phorbol esters increases the AQP1 cationic current by a mechanism that is dependent on phosphorylation of Thr157 and Thr239 residues45 located in loop D and in the C-terminus, respectively. In Site 2, an Arg residue in the carboxyl terminus is conserved (243 in bAQP1; 261 in rAQP4). In continuing work, it will be of interest to determine whether any of the arylsulfonamide-related derivatives can be found to act at Site 2, at which possible modulatory effects on the water or ion fluxes might be identified. Such tools will be valuable for addressing the physiological purpose of dual channel function in the aquaporins that are both ion and water channels, such as AQP0,49,50 AQP143,44 and AQP6,51 and potentially other AQP ion channels yet to be identified.
Calls for the development of aquaporin blockers have been driven by recognition of the protective effects of genetic aquaporin deficiency against pathological fluid accumulation and disease processes in mouse models. AQPs have an obvious role in mediating fluid homeostasis, but also have been associated with diverse other processes including angiogenesis, cell migration, development, neuropathological disease, and cancer, through mechanisms that are not yet understood. Examples of two areas of particular interest for investigating translational value of the AqB compounds include treating brain oedema, and as a possible adjunct for novel strategies of cancer treatment.
Brain oedema is a potentially fatal consequence of head injury, brain tumour, stroke, and infections such as meningitis, all of which cause brain swelling due to excess fluid accumulation.29,52-54 Cerebral oedema is the leading cause of mortality and morbidity associated with head trauma and stroke.55 Current treatments include neurosurgical decompression and administration of hyperosmolar agents to extract brain water osmotically.56,57 AQP4 is postulated to play a critical role in the formation of brain oedema after stroke, injury or water intoxication. Decreased AQP4 expression in mammals can reduce the rate of pathological fluid accumulation in the brain, and AQP4 deficient mice show improved outcomes in models of brain oedema. AQP4-null mice demonstrated reduced brain water content and astrocytic swelling, and improved neurological outcomes and survival after ischemic stroke, as compared with wild-type mice.53 Injection of Streptococcus pneumoniae causes meningitis-induced brain oedema; AQP4 null mice had significantly lower intracranial pressure and brain water accumulation, and a higher survival (80% compared to 0% at 60h) as compared with wild type mice.54 The reduced brain oedema seen in the AQP4-deficient mice highlights this channel as a potential target for drug therapy.
AQP1 is also an important therapeutic target in the brain. Imbalances between cerebral spinal fluid production and absorption can result in hydrocephalus, increased intracranial pressure, and mental retardation. Potential AQP1 blockers could have a protective effect in related disorders. In a rodent model of focal brain injury, AQP1-deficient mice demonstrated a five-fold reduction in osmotically driven water flux across the choroid epithelium, a reduction in intracranial pressure and better survival rates than in the wild-type mice.58
The upregulation of the water channel Aquaporin-1 (AQP1) is a unique feature of glioblastoma and certain other cancer types (breast and colon cancers), but the role of AQP1 in cancer has been difficult to evaluate, given the relative lack of available pharmacological agents.59,60 Glioblastomas are invasive brain tumours that result from transformation of glial cells. In these rapidly lethal tumours the level of Aquaporin-1 expression has been found to correlate with the aggressiveness of the disease, although the role of Aquaporin-1 in the disease process is not known. The age distribution in the world population of humans with brain tumours is bimodal, with peak incidences in children and adults between ages 45 to 70, and the highest rates seen in descendants of white European ethnic groups. Glioblastoma multiforme is the most common, malignant and aggressive form of brain tumour in adults, and accounts for 75-80% of primary brain gliomas as the 12th leading cause of cancer-related death in men. With glioblastoma multiforme, most patients develop clinical symptoms over a short time span (three months) and die within 8-18 months of diagnosis.61 Less than half of the patients diagnosed with glioblastoma live for 6 months, and only 3% survive for two years. The incidence rate of glioblastoma per year is 3.32 in males and 2.24 in females per 100,000; of these cases, the majority are de novo glioblastomas, and rarely involve progression from low grade gliomas (5%).62
It is possible that block of AQP channels could slow the proliferation and migration of glioma cells. The upregulation of AQP1 expression in glioblastoma enables cancer cell volume regulation and promotes survival, proliferation, and migration.63 The suggested role of AQP1 in the pathology of aggressive glioblastoma has been that the water channel provides for a regulated decrease in tumour cell volume which confers increased invasiveness of them through the restricted extracellular space of the central nervous system. However, an in depth consideration of the available data for AQPs in cancer (below) shows that more than just water channel function is involved; specific classes of AQPs are associated with the different classes of tumours, and the transformative effects of AQP overexpression on morphology, growth and proliferation cannot be mimicked by substitution of other classes of water channels.
In primary glioblastoma multiformae tumours, receptors and transporters that are involved in glutamate and GABA neurotransmission are downregulated as compared with normal temporal lobe human brain tissue, but in contrast, AQP1 expression is upregulated.64 Glioma biopsies consistently show upregulation of AQP1, though the expression is lost after the malignant cells are isolated and propagated in culture. In glioblastoma multiforme, upregulated AQP1 expression is proposed to contribute to oedema formation and cell motility. AQP1 expression was found to be strongly upregulated in human glial tumours available in the UCSF Neurosurgery Tissue Bank (36 samples), and might contribute to the formation of oedema associated with malignant glial tumours.65,66
In contrast, AQP4 does not have an obvious link in glioblastoma. AQP4 is normally expressed in the perivascular endfeet of astrocytes; however in gliomas, the normal localisation is lost and the channels are dispersed over the cell surface. Levels of expression of AQP4 in Grade I to IV gliomas (World Health Organisation scale) were associated with the extent of oedema formation, but showed no correlation with patient survival.67 While introduction of AQP4 did increase water permeability as expected, its additional effect was to enhance cell adhesion. The increase in AQP4 did not alter growth or migration.63
In human lung cancers, AQP1 is overexpressed in adeno- and bronchoalveolar carcinomas, but not in squamous cell carcinoma or in normal lung tissue.68 Over- expression of AQP1 in NIH-3T3 cells induces phenotypic changes characteristic of transformation, including cell proliferation and anchorage-independent growth in soft agar. Upregulation of AQP5 expression correlates with an increased rate of recurrence of non-small cell lung cancer, and a decreased chance of disease-free survival. Transfection of cell lines with wild type AQP5 altered the cell morphology to a fibroblastic phenotype, accompanied by the loss of cell-cell contacts, polarity, and epithelial markers, and the gain of mesenchymal cell markers; whereas mutant constructs lacking trafficking or phosphorylation signal sites did not cause these alterations.69 Overexpression of human AQP5 induces phenotypic changes that are characteristic of transformation of cells in vitro and in vivo. An increase of protein-kinase\-A mediated phosphorylation of AQP5 was observed in tumour cells and associated with increased proliferation.70
In colorectal carcinomas, microarray profiling has shown that AQP1 can be considered to be a validated gene that discriminates colorectal cancer cells from adenomas and normal tissue.71,72 Overexpression of human AQP5 activates phosphorylation signaling cascades and induces proliferation in a colon cancer cell line. The effect is not replicated by the overexpression of other aquaporins AQP1 and AQP3,73 indicating that the class of AQP linked to the pathogenic state depends on the type of tissue involved. AQP1 and AQP5 are expressed in colorectal cancer, from the early-stage of dysplasia through the late stages of colon cancer development, and in the metastatic lesions found in liver.74 The implied role for AQP expression in tumorigenesis at early stages makes AQP an attractive target for potential therapeutic strategies.
In primary renal carcinomas, expression of AQP1 was significantly higher in clear-cell and papillary forms, even at early stages before symptomatic presentation, as compared with the normal levels of expression of this protein in kidney tissue samples. It is interesting to note that the addition of the AQP1 parameter to the University of California Los Angeles Integrated Staging System (UISS) prognostic score improved the accuracy of predictions of both cancer death and recurrence for all patient cohorts for clear-cell renal carcinoma.75 In mammary carcinomas, tumour cell membranes show significant expression of AQP1 that is not seen in the normal epithelial cells.76
Together these results suggest that the cancer-enhancing activity of AQPs is more than simply creating pathways for water flow, but that additional distinguishing features (such as capacity for linkages of AQPs into protein-protein signaling complexes, or regulation by intracellular signaling pathways) are also important. The role of AQPs in human cancer is emerging as an area of intense research interest. Analyses of AQP knockout mice have confirmed the expected functions of AQPs in epithelial fluid transport. However, some surprising roles for AQPs have emerged, in that AQPs have been found to promote cell migration, tumour angiogenesis, tumour extravasation and metastasis, to alter fat metabolism and more, by mechanisms that remain unknown.60 Recognizing that AQP1 is a water channel and, under permissive conditions, also a cGMP-gated cation channel,43,45,47 evidence in various tissues for a coupling of the cGMP signaling cascade to a physiological outcome that might involve AQP1 dual ion-and-water channel functions is of interest.3,14
It appears that the call for development of pharmacological agents for aquaporins is being answered, and the field is poised for exciting progress in basic science and medicine. With the new compounds being identified, it is possible to finally begin testing for a role of AQP blockers in mitigating pathologies such as brain oedema, glioblastoma, breast and colon cancers, and many other clinically important disorders.
Supported by National Institutes of Health RO1 GM059986 and the University of Adelaide School of Molecular & Biomedical Science.
1. Agre P. Aquaporin water channels (Nobel Lecture). Angew Chem. Int. Ed. Engl. 2004; 43: 4278-90.
2. Musa-Aziz R, Chen LM, Pelletier MF, Boron WF. Relative CO2/NH3 selectivities of AQP1, AQP4, AQP5, AmtB, and RhAG. Proc. Natl. Acad. Sci. USA 2009; 106:5406-11.
3. Yool AJ. Aquaporins: multiple roles in the central nervous system. Neuroscientist 2007; 13: 470-85.
4. Preston GM, Agre P. Isolation of the cDNA for erythrocyte integral membrane protein of 28 kilodaltons: member of an ancient channel family. Proc. Natl. Acad. Sci. USA 1991; 88: 11110-4.
5. Benga G, Popescu O, Pop VI, Holmes RP. p-(Chloromercuri)benzenesulfonate binding by membrane proteins and the inhibition of water transport in human erythrocytes. Biochemistry 1986; 25: 1535-8.
6. Stamer WD, Snyder RW, Smith BL, Agre P, Regan JW. Localization of aquaporin CHIP in the human eye: implications in the pathogenesis of glaucoma and other disorders of ocular fluid balance. Invest. Ophthalmol. Vis. Sci. 1994; 35:3867-72.
7. Au CG, Cooper ST, Lo HP, et al. Expression of aquaporin 1 in human cardiac and skeletal muscle. J. Mol. Cell. Cardiol. 2004; 36: 655-62.
8. Nielsen S, Smith BL, Christensen EI, Agre P. Distribution of the aquaporin CHIP in secretory and resorptive epithelia and capillary endothelia. Proc. Natl. Acad. Sci. USA 1993; 90: 7275-9.
9. Boassa D, Stamer WD, Yool AJ. Ion channel function of aquaporin-1 natively expressed in choroid plexus. J. Neurosci .2006; 26: 7811-9.
10. Johansson PA, Dziegielewska KM, Ek CJ, et al. Aquaporin-1 in the choroid plexuses of developing mammalian brain. Cell Tissue Res. 2005; 322: 353-64.
11. Jung JS, Preston GM, Smith BL, Guggino WB, Agre P. Molecular structure of the water channel through aquaporin CHIP. The hourglass model. J. Bio. Chem. 1994; 269: 14648-54.
12. Sui H, Han BG, Lee JK, Walian P, Jap BK. Structural basis of water-specific transport through the AQP1 water channel. Nature 2001; 414: 872-8.
13. Murata K, Mitsuoka K, Hirai T, et al. Structural determinants of water permeation through aquaporin-1. Nature 2000; 407: 599-605.
14. Yool AJ. Functional domains of aquaporin-1: keys to physiology, and targets for drug discovery. Cur. Pharm. Des. 2007; 13:3212-21.
15. Zador Z, Bloch O, Yao X, Manley GT. Aquaporins: role in cerebral edema and brain water balance. Prog. Brain Res. 2007; 161: 185-94.
16. Preston GM, Jung JS, Guggino WB, Agre P. The mercury-sensitive residue at cysteine 189 in the CHIP28 water channel. J. Bio. Chem. 1993; 268: 17-20.
17. Niemietz CM, Tyerman SD. New potent inhibitors of aquaporins: silver and gold compounds inhibit aquaporins of plant and human origin. FEBS Lett. 2002; 531: 443-7.
18. Ma B, Xiang Y, Mu SM, Li T, Yu HM, Li XJ. Effects of acetazolamide and anordiol on osmotic water permeability in AQP1-cRNA injected Xenopus oocyte. Acta Pharmacol. Sin. 2004; 25: 90-7.
19. Gao J, Wang X, Chang Y, et al. Acetazolamide inhibits osmotic water permeability by interaction with aquaporin-1. Anal. Biochem. 2006; 350: 165-70.
20. Brooks HL, Regan JW, Yool AJ. Inhibition of aquaporin-1 water permeability by tetraethylammonium: involvement of the loop E pore region. Mol. Pharmacol. 2000; 57: 1021-6.
21. Yool A, Brokl O, Pannabecker T, Dantzler W, Stamer W. Tetraethylammonium block of water flux in Aquaporin-1 channels expressed in kidney thin limbs of Henle's loop and a kidney-derived cell line. BMC physiol. 2002; 2: 4.
22. Detmers FJ, de Groot BL, Muller EM, et al. Quaternary ammonium compounds as water channel blockers. Specificity, potency, and site of action. J. Biol. Chem. 2006; 281: 14207-14.
23. Yang B, Kim JK, Verkman AS. Comparative efficacy of HgCl2 with candidate aquaporin-1 inhibitors DMSO, gold, TEA+ and acetazolamide. FEBS Lett. 2006; 580: 6679-84.
24. Kuppers E, Gleiser C, Brito V, et al. AQP4 expression in striatal primary cultures is regulated by dopamine--implications for proliferation of astrocytes. Eur. J. Neurosci. 2008; 28: 2173-82.
25. Watson KJ, Kim I, Baquero AF, Burks CA, Liu L, Gilbertson TA. Expression of aquaporin water channels in rat taste buds. Chem. Senses 2007; 32: 411-21.
26. Migliati E, Meurice N, Dubois P, et al. Inhibition of Aquaporin-1 and Aquaporin-4 water permeability by a derivative of the loop diuretic bumetanide acting at a internal pore-occluding binding site. Mol. Pharmacol. 2009. doi:10.1124/mol.108.053744.
27. Amiry-Moghaddam M, Ottersen OP. The molecular basis of water transport in the brain. Nat. Rev. Neurosci. 2003; 4: 991-1001.
28. Jung JS, Bhat RV, Preston GM, Guggino WB, Baraban JM, Agre P. Molecular characterization of an aquaporin cDNA from brain: candidate osmoreceptor and regulator of water balance. Proc. Natl. Acad. Sci. USA 1994; 91: 13052-6.
29. Amiry-Moghaddam M, Otsuka T, Hurn PD, et al. An a-syntrophin-dependent pool of AQP4 in astroglial end-feet confers bidirectional water flow between blood and brain. Proc. Natl. Acad. Sci. USA 2003; 100: 2106-11.
30. Ma T, Yang B, Gillespie A, Carlson EJ, Epstein CJ, Verkman AS. Generation and phenotype of a transgenic knockout mouse lacking the mercurial-insensitive water channel aquaporin-4. J. Clin. Invest. 1997; 100: 957-62.
31. Yukutake Y, Tsuji S, Hirano Y, et al. Mercury chloride decreases the water permeability of aquaporin-4-reconstituted proteoliposomes. Biol. Cell 2008; 100: 355-63.
32. Huber VJ, Tsujita M, Yamazaki M, Sakimura K, Nakada T. Identification of arylsulfonamides as Aquaporin 4 inhibitors. Bioorg. Med. Chem. Lett. 2007; 17:1270-3.
33. Huber VJ, Tsujita M, Kwee IL, Nakada T. Inhibition of Aquaporin 4 by antiepileptic drugs. Bioorg. Med. Chem. 2008.
34. Haas M, McManus TJ. Bumetanide inhibits Na+-K+-2Cl− co-transport at a chloride site. Am. J. Physiol. 1983; 245: C235-40.
35. O'Donnell ME, Tran L, Lam TI, Liu XB, Anderson SE. Bumetanide inhibition of the blood-brain barrier Na+-K+-Cl− cotransporter reduces edema formation in the rat middle cerebral artery occlusion model of stroke. J. Cereb. Blood Flow Metab. 2004; 24:1046-56.
36. Lam TI, Anderson SE, Glaser N, O'Donnell ME. Bumetanide reduces cerebral edema formation in rats with diabetic ketoacidosis. Diabetes 2005; 54: 510-6.
37. Yan Y, Dempsey RJ, Sun D. Na+-K+-Cl− cotransporter in rat focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2001; 21: 711-21.
38. Yan Y, Dempsey RJ, Flemmer A, Forbush B, Sun D. Inhibition of Na+-K+-Cl− cotransporter during focal cerebral ischemia decreases edema and neuronal damage. Brain Res. 2003; 961: 22-31.
39. Yu J, Yool AJ, Schulten K, Tajkhorshid E. Mechanism of gating and ion conductivity of a possible tetrameric pore in aquaporin-1. Structure 2006; 14: 1411-23.
40. Zelenina M, Zelenin S, Bondar AA, Brismar H, Aperia A. Water permeability of aquaporin-4 is decreased by protein kinase C and dopamine. Am. J. Physiol. 2002; 283: F309-18.
41. Fotiadis D, Suda K, Tittmann P, et al. Identification and structure of a putative Ca2+-binding domain at the C terminus of AQP1. J. Mol. Biol. 2002; 318: 1381-94.
42. Tornroth-Horsefield S, Wang Y, Hedfalk K, et al. Structural mechanism of plant aquaporin gating. Nature 2006; 439: 688-94.
43. Anthony TL, Brooks HL, Boassa D, et al. Cloned human aquaporin-1 is a cyclic GMP-gated ion channel. Mol. Pharmacol. 2000; 57: 576-88.
44. Yool AJ, Stamer WD, Regan JW. Forskolin stimulation of water and cation permeability in aquaporin 1 water channels. Science 1996; 273: 1216-8.
45. Zhang W, Zitron E, Homme M, et al. Aquaporin-1 channel function is positively regulated by protein kinase C. J. Biol. Chem. 2007; 282: 20933-40.
46. Yool AJ, Weinstein AM. New roles for old holes: Ion channel function in aquaporin-1. NIPS 2002; 17: 68-72.
47. Boassa D, Yool AJ. A fascinating tail: cGMP activation of aquaporin-1 ion channels. Trends Pharmacol. Sci. 2002; 23: 558-62.
48. Boassa D, Yool AJ. Single amino acids in the carboxyl terminal domain of aquaporin-1 contribute to cGMP-dependent ion channel activation. BMC Physiol. 2003; 3: 12.
49. Ehring GR, Hall JE. Single channel properties of lens MIP 28 reconstituted into planar lipid bilayers. Proc. West. Pharmacol. 1988; 31:251-3.
50. Zampighi GA, Hall JE, Kreman M. Purified lens junctional protein forms channels in planar lipid films. Proc. Natl. Acad. Sci. USA 1985; 82: 8468-72.
51. Yasui M, Hazama A, Kwon TH, Nielsen S, Guggino WB, Agre P. Rapid gating and anion permeability of an intracellular aquaporin. Nature 1999; 402: 184-7.
52. Amiry-Moghaddam M, Frydenlund DS, Ottersen OP. Anchoring of aquaporin-4 in brain: molecular mechanisms and implications for the physiology and pathophysiology of water transport. Neuroscience 2004; 129: 999-1010.
53. Manley GT, Fujimura M, Ma T, et al. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat. Med. 2000; 6: 159-63.
54. Papadopoulos MC, Verkman AS. Aquaporin-4 gene disruption in mice reduces brain swelling and mortality in pneumococcal meningitis. J. Biol. Chem. 2005; 280: 13906-12.
55. Klatzo I. Evolution of brain edema concepts. Acta Neurochir. Suppl. (Wien) 1994; 60: 3-6.
56. Ogden AT, Mayer SA, Connolly ES, Jr. Hyperosmolar agents in neurosurgical practice: the evolving role of hypertonic saline. Neurosurgery 2005; 57: 207-15; discussion -15.
57. Subramaniam S, Hill MD. Massive cerebral infarction. Neurologist 2005; 11: 150-60.
58. Oshio K, Watanabe H, Song Y, Verkman AS, Manley GT. Reduced cerebrospinal fluid production and intracranial pressure in mice lacking choroid plexus water channel Aquaporin-1. FASEB J 2005; 19: 76-8.
59. Jeyaseelan K, Sepramaniam S, Armugam A, Wintour EM. Aquaporins: a promising target for drug development. Expert. Opin. Ther. Targets 2006; 10: 889-909.
60. Verkman AS, Hara-Chikuma M, Papadopoulos MC. Aquaporins — new players in cancer biology. J. Mol. Med. 2008; 86: 523-9.
61. Burger PC, Green SB. Patient age, histologic features, and length of survival in patients with glioblastoma multiforme. Cancer 1987; 59: 1617-25.
62. Ohgaki H, Dessen P, Jourde B, et al. Genetic pathways to glioblastoma: a population-based study. Cancer Res 2004; 64: 6892-9.
63. McCoy E, Sontheimer H. Expression and function of water channels (aquaporins) in migrating malignant astrocytes. Glia 2007; 55: 1034-43.
64. Markert JM, Fuller CM, Gillespie GY, et al. Differential gene expression profiling in human brain tumors. Physiol. Genomics 2001; 5: 21-33.
65. Oshio K, Binder DK, Bollen A, Verkman AS, Berger MS, Manley GT. Aquaporin-1 expression in human glial tumors suggests a potential novel therapeutic target for tumor-associated edema. Acta Neurochir. Suppl. 2003; 86: 499-502.
66. Saadoun S, Papadopoulos MC, Davies DC, Bell BA, Krishna S. Increased aquaporin 1 water channel expression in human brain tumours. Br. J. Cancer 2002; 87: 621-3.
67. Warth A, Simon P, Capper D, et al. Expression pattern of the water channel aquaporin-4 in human gliomas is associated with blood-brain barrier disturbance but not with patient survival. J. Neurosci. Res. 2007; 85: 1336-46.
68. Hoque MO, Soria JC, Woo J, et al. Aquaporin 1 is overexpressed in lung cancer and stimulates NIH-3T3 cell proliferation and anchorage-independent growth. Am. J. Pathol. 2006; 168: 1345-53.
69. Chae YK, Woo J, Kim MJ, et al. Expression of aquaporin 5 (AQP5) promotes tumor invasion in human non small cell lung cancer. PLoS ONE 2008; 3: e2162. doi:10.1371/journal.pone.0002162
70. Woo J, Lee J, Chae YK, et al. Overexpression of AQP5, a putative oncogene, promotes cell growth and transformation. Cancer Lett. 2008; 264: 54-62.
71. Galamb O, Gyorffy B, Sipos F, et al. Inflammation, adenoma and cancer: objective classification of colon biopsy specimens with gene expression signature. Dis. Markers 2008; 25: 1-16.
72. Galamb O, Sipos F, Solymosi N, et al. Diagnostic mRNA expression patterns of inflamed, benign, and malignant colorectal biopsy specimen and their correlation with peripheral blood results. Cancer Epidemiol. Biomarkers Prev. 2008; 17: 2835-45.
73. Kang SK, Chae YK, Woo J, et al. Role of human aquaporin 5 in colorectal carcinogenesis. Am. J. Pathol. 2008; 173: 518-25.
74. Moon C, Soria JC, Jang SJ, et al. Involvement of aquaporins in colorectal carcinogenesis. Oncogene 2003; 22: 6699-703.
75. Huang Y, Murakami T, Sano F, et al. Expression of aquaporin 1 in primary renal tumors: A prognostic indicator for clear-cell renal cell carcinoma. Eur. Urol. 2008. doi:10.1016/j.eururo.2008.10.014.
76. Endo M, Jain RK, Witwer B, Brown D. Water channel (aquaporin 1) expression and distribution in mammary carcinomas and glioblastomas. Microvasc. Res. 1999; 58: 89-98.