At the start of tetanic contractions, mammalian skeletal muscle fibres are often excited by high frequency motorneuron double discharges, of 200 Hz or more (Desmedt & Godauxa, 1977). Recently, Cheng et al., 2013 reported that the presence of an initial 200 Hz doublet action potential can result in a 100% increase in peak tetanic Ca2+ compared to control Ca2+ transients elicited at a stimulation rate of 70Hz alone, there was also a faster rise in force (Cheng et al., 2013). However, Cheng et al. tracked Ca2+ release with only ms temporal resolution, leaving the evolution of the Ca2+ transient that underlies the rapidly rising force response unknown. In this study, we imaged Ca2+ release with μs resolution using a Zeiss 5 Live during doublet action potential stimulation in conjunction with the fast, low affinity Ca2+ indicator Mag-Fluo-4.
Mice were killed by cervical dislocation, and the interosseous muscles removed and placed in a collagenase II digestion solution (mg/ml) for 30 min. The muscles were then gently triturated to produce single fibres. Fibres were loaded with Mag-Fluo-4 (5μM for 15 min), and placed in a solution bath containing a HEPES based Kreb’s Ringer, and the myosin inhibitor BTS (100 μM) to prevent fibre movement. Fast fibres were selected based on the shape of elicited tetanic Ca2+ transients (Calderon, Bolanos & Caputo, 2011). In control measurements, fibres were activated by 10 action potentials at 120 Hz using platinum electrodes. Stimulation during the doublet measurements was similar with the exception that the first 2 action potentials were at 200 Hz. Ca2+ fluorescence was captured at ∼10 kHz using a Zeiss 5 Live confocal microscope in linescan mode. Changes in the progression of the Ca2+ response was quantitated by estimating the lowest Ca2+ fluorescence found after each Ca2+ spike, and normalising the value to the fluorescence nadir after the 6th response.
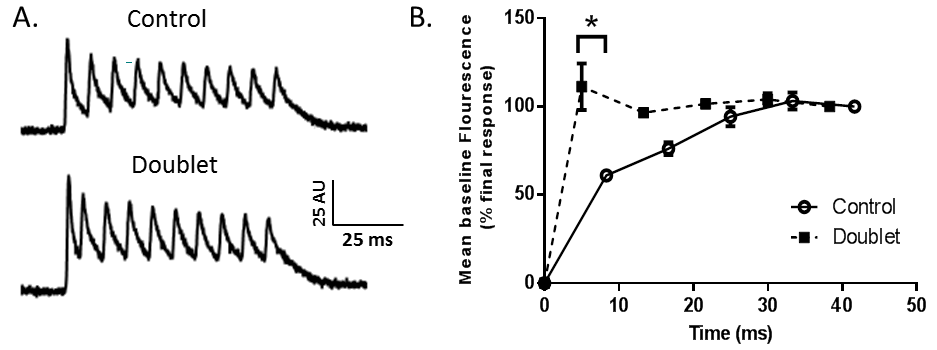
A. Effects of control and doublet stimulation on Mag-fluo-4 fluorescence (AU: arbitrary units). B. Changes in the minimum fluorescence value between Ca2+ transient spikes before (time 0) and after activation under control conditions or with an initial 120 Hz doublet (*P<0.05).
The presence of an initial 200 Hz doublet action potential did not significantly alter the amplitudes of the Ca2+ spikes during the transient. However, after doublet stimulation, the minimum fluorescence value between Ca2+ transient spikes rose to a stable, maximal level after the first response that was approximately 1.8 times greater than the basal Ca2+ fluorescence after the first response under control conditions (control: 61.06 ± 2.82% of 6th response; doublet: 111.27 ± 13.19% of 6th response) (Figure). Furthermore, in controls, the minimum fluorescence value between Ca2+ transient spikes did not reach the maximum normalised value until 16 - 25 ms after initial activation, compared to only 5 ms in fibres exposed to doublet stimulation.
These results indicate that doublet stimulation rapidly increases the minimum fluorescence value between Ca2+ transient spikes in fast twitch muscle fibres. Doublet activation may lead to more rapid saturation of cytosolic Ca2+ binding sites and therefore, faster initiation of cross-bridge cycling in fast skeletal muscle fibres.
Desmedt JE, Godaux E. (1977) Ballistic contractions in man: characteristic recruitment pattern of single motor units of the tibialis anterior muscle. Journal of Physiology 264: 673-93.
Cheng AJ, Place N, Bruton JD, Holmberg HC, Westerblad H. (2013) Doublet discharge stimulation increases sarcoplasmic reticulum Ca2+ release and improves performance during fatiguing contractions in mouse muscle fibres. Journal of Physiology 591: 3739-48.
Calderon JC, Bolanos P, Caputo C. (2011) Kinetic changes in tetanic Ca2+ transients in enzymatically dissociated muscle fibres under repetitive stimulation. Journal of Physiology 589: 5269-83.