Depletion of calcium (Ca2+) from intracellular stores (endoplasmic reticulum, ER), triggers Ca2+ entry across the plasma membrane, a process known as capacitative Ca2+ entry or store-operated Ca2+ entry (SOCE) (Parekh & Penner, 1997). SOCE is important for intracellular Ca2+ regulation and is mediated by the interaction between STIM1 (stromal interaction molecule-1), which functions as the Ca2+ sensor in the ER (Liou et al., 2005), and Ca2+ permeable Orai1 channels in the external membrane (Vig et al., 2006). SOCE in skeletal muscle fibres (Kurebayashi & Ogawa, 2001) is also mediated by interaction between STIM1 in the sarcoplasmic reticulum (SR) and Orai1 channels in transverse tubules (TT) (Lyfenko & Dirksen, 2008).
The question. As this mechanism is likely the primary mechanism of Ca2+ entry in muscle during repetitive activity, it has been proposed that SOCE is important to prevents/delays fatigue (Wei-Lapierre et al., 2013). Despite its importance for proper muscle function, the subcellular sites for SOCE in skeletal fibres have not been unequivocally identified.
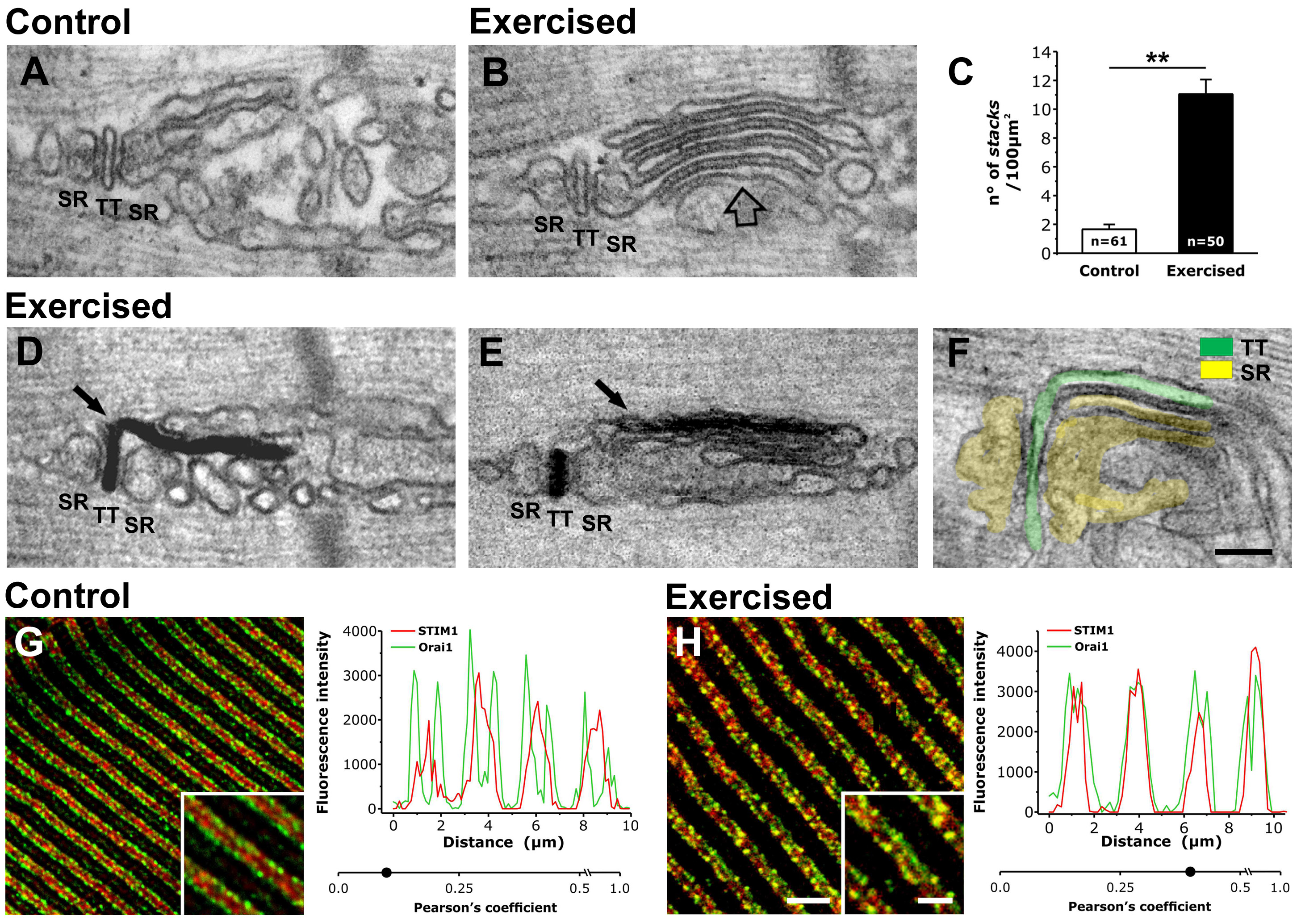
Main findings. Using electron microscopy we show that prolonged muscle activity (treadmill running in mice) drives the formation of previously unidentified intracellular junctions between SR and extensions of the TT membrane at the I band, between Ca2+-release units (CRUs, the sites of excitation-contraction coupling) and Z-line. Using immunohistochemistry and immunogold labeling we also demonstrate that these newly formed junctions contain the molecular machinery known to mediate SOCE in muscle: STIM1 Ca2+ sensor proteins in the SR, already present in the I band in control conditions, and Ca2+-permeable Orai1 channels, which move into the I band with TTs during prolonged muscle activity.
The Figure Panels A-B: show that following exercise, internal membranes at the I band rearrange into stacks of flat cisternae (empty arrow in B). C: Incidence of membrane stacks (as the one shown in B) is much higher in exercised muscles. D-F: Following exercise, TTs (stained black with ferrocyanide-precipitate in D and E) grows into the I band and become part of stacks of cisternae. G and H) Colocalization between STIM1 and Orai1 is low in control samples, but increases significantly following exercise. SR-TT-SR labels a triad (the site of excitation-contraction coupling in muscle). Data in C are shown as mean ± SEM: **p<0.01. Scale bars: A-F, 0.1 μm; G and H, 2.5 μm (insets: 1 μm).
Conclusions. The activity-dependent formation of these unique SR-TT junctions reflects a striking and unexpected remodeling of the existing sarcotubular system at the I band of the sarcomere. We here name these junctions Ca2+ Entry Units (CEUs), and propose that these structural entities are the preferential site for SOCE, at least during repetitive activity, representing the ideal pathway to rapidly recover Ca2+ ions from the extracellular space and, hence, important to prevent/delay fatigue.
Additional implications. As a) reduced SOCE activity also contributes to muscle dysfunction in ageing (Zhao et al., 2008), and b) mutations in STIM1 and Orai1 are linked to Tubular Aggregate Myopathy (Nesin et al., 2014), our findings could also have important implications for the understanding of muscular dysfunction in different physio-pathological conditions.
Kurebayashi N & Ogawa Y. (2001). Depletion of Ca2+ in the sarcoplasmic reticulum stimulates Ca2+ entry into mouse skeletal muscle fibres. J Physiol 533, 185-99.
Liou J, Kim ML, Heo WD, Jones JT, Myers JW, Ferrell JE Jr, Meyer T. (2005). STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr Biol 15, 1235-41.
Lyfenko AD & Dirksen RT. (2008). Differential dependence of store-operated and excitation-coupled Ca2+ entry in skeletal muscle on STIM1 and Orai1. J Physiol 586, 4815-24.
Nesin V, Wiley G, Kousi M, Ong EC, Lehmann T, Nicholl DJ, Suri M, Shahrizaila N, Katsanis N, Gaffney PM, Wierenga KJ, Tsiokas L. (2014). Activating mutations in STIM1 and ORAI1 cause overlapping syndromes of tubular myopathy and congenital miosis. Proc Natl Acad Sci USA 111, 4197-202.
Parekh AB & Penner R. (1997). Store depletion and calcium influx. Physiol Rev 77, 901-30.
Vig M, Peinelt C, Beck A, Koomoa DL, Rabah D, Koblan-Huberson M, Kraft S, Turner H, Fleig A, Penner R, Kinet JP. (2006). CRACM1 is a plasma membrane protein essential for store-operated Ca2+ entry. Science 312, 1220-3.
Wei-Lapierre L, Carrell EM, Boncompagni S, Protasi F, Dirksen RT. (2013). Orai1-dependent calcium entry promotes skeletal muscle growth and limits fatigue. Nat Commun 4, 2805.
Zhao X1, Weisleder N, Thornton A, Oppong Y, Campbell R, Ma J, Brotto M. (2008). Compromised store-operated Ca2+ entry in aged skeletal muscle. Aging Cell 7, 561-8.