Syntaxins are single-pass membrane-anchored proteins, which in their classic role form part of SNARE complex mediating vesicular fusion events. The first syntaxin to be characterized in this role was syntaxin 1A – a T-SNARE facilitating neurotransmitter-containing vesicular fusion at neuronal synapses. Another well-known role of syntaxin 1A is its functional interaction with a wide range of membrane transporters and channels, including the GABA, dopamine, and norepinephrine transporters of the Neurotransmitter Sodium Symporter (NSS)/SLC6 family. Close homologues to these NSS neurotransmitter re-uptake transporters are two kidney epithelial transporters of the SLC6 family B0AT1 and B0AT3, also characterized by their association with collectrin, another single-pass membrane protein which act as a molecular chaperone to the transporters. Collectrin has further been implicated in renal aminoacidurias, hypertension, insulin exocytosis and β-cell proliferation in the pancreas.
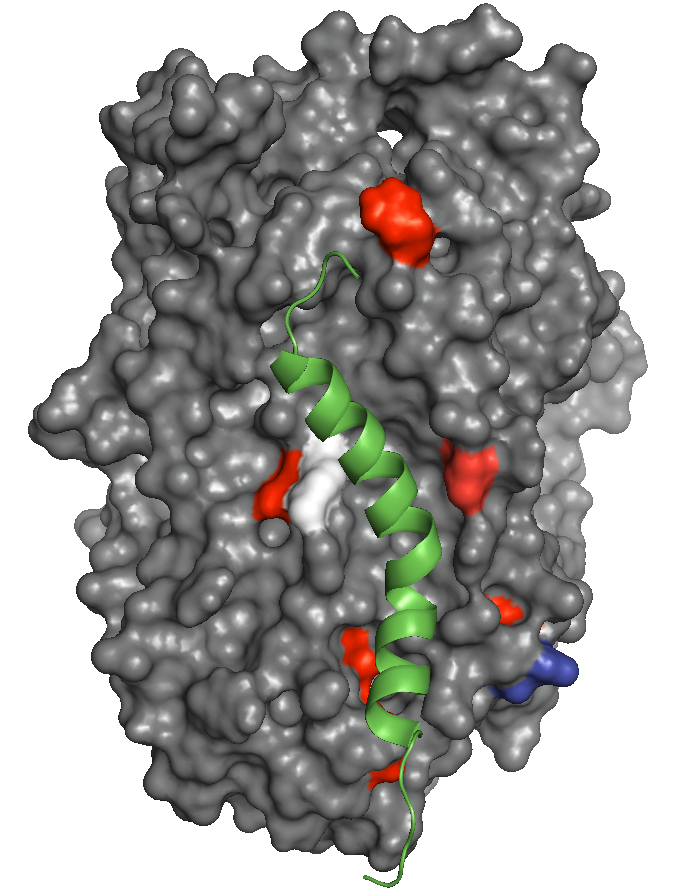
We show that sytaxin 1A and the kidney specific syntaxin 3 also interact with B0AT1, and collectrin with the GABA transporter. Consistent with their role in regulating neurotransmitter transporters, syntaxins inhibit B0AT1 trafficking to the plasma membrane, whilst the effect of collectrin is to stimulate trafficking. We also investigated the possibility that the binding of these unrelated ancillary proteins represents a common competitive binding site conserved across SLC6 transporters. In addition, and unexpectedly, we also discovered collectrin is essential for the catalytic activation and functional complex formation of both B0AT1 and B0AT3. Sequence alignment, structural considerations and mutational analysis suggest that amino acid residues clustering around a cavity formed by trans-membrane helices 5 and 7 are vital for collectrin-mediated membrane stabilization and functional activation. These findings demonstrate that B0AT1 is a heteromeric transporter forming a functional complex with collectrin and possibly competing with syntaxin proteins at a common SLC6 binding site.